


%20-%20Made%20with%20PosterMyWall.jpg)
 ISBN: 978-618-86274-5-1
ISBN: 978-618-86274-5-1
 ISBN: 978-618-86274-4-4
ISBN: 978-618-86274-4-4
%20-%20Made%20with%20PosterMyWall.jpg)




Μπορείτε να το κατεβάσετε από εδώ:
%20-%20Made%20with%20PosterMyWall.jpg) ISBN: 9-786180--052817
ISBN: 9-786180--052817 Μπορείτε να το κατεβάσετε από εδώ:
Μπορείτε να το κατεβάσετε από εδώ:
 ISBN:978-618-86274-3-7
ISBN:978-618-86274-3-7 Το πλήρες e-book σε pdf μπορείτε να το κατεβάζετε δωρεάν από το παρακάτω link:
Το πλήρες e-book σε pdf μπορείτε να το κατεβάζετε δωρεάν από το παρακάτω link:

ISBN:978-618-00-5245-9
 Το πλήρες e-book σε pdf μπορείτε να το κατεβάζετε δωρεάν από το παρακάτω link:
Το πλήρες e-book σε pdf μπορείτε να το κατεβάζετε δωρεάν από το παρακάτω link:

ISBN:978-618-86274-2-0
Η αλπικού τύπου Δρακόλιμνη στο Βελούχι
Μια επιστημονική προσέγγιση με την χρήση των GIS
Μπακολίτσας ΚωνσταντίνοςΒιολόγος - Εκπαιδευτής GIS
Το πλήρες e-book σε pdf μπορείτε να το κατεβάζετε δωρεάν από το παρακάτω link:https://drive.google.com/file/d/10xpDIQsPJIbiUMnIqDTTWW-tmn5ajh3U/view?usp=sharing
Βιογραφικό του συγγραφέα:
Επικοινωνία: sv1cid@gmail.com
%20-%20Made%20with%20PosterMyWall.jpg)



ISBN:978-618-00-5245-9



Μια επιστημονική προσέγγιση με την χρήση των GIS
12. Τεχνικές αλυσιδωτής αντίδρασης πολυμεράσης: Αρχή μεθόδων, διδακτικά σχήματα, εφαρμογές.
Η αλυσιδωτή αντίδραση πολυμεράσης (PCR) είναι μία τεχνική στη βιοχημεία και στη μοριακή βιολογία [1] για ενζυμική αντιγραφή του DNA χωρίς χρησιμοποίηση ζωντανών οργανισμών , όπως η E. coli ή η ζύμη . Όπως στην ενίσχυση με τη χρησιμοποίηση ζωντανών οργανισμών, η τεχνική επιτρέπει ένα μικρό ποσό DNA για να ενισχυθεί εκθετικά. Όπως η PCR είναι in vitro τεχνική, μπορεί να εκτελεσθεί χωρίς περιορισμούς στη μορφή του DNA, και μπορεί να είναι εκτενώς τροποποιημένη για να εκτελέσει μια ευρεία σειρά γενετικών χειρισμών.[2]
Η PCR χρησιμοποιείται συνήθως στα ιατρικά και βιολογικά ερευνητικά εργαστήρια για ποικίλους στόχους, όπως η ανίχνευση κληρονομικών ασθενειών, ο προσδιορισμός γενετικών δακτυλικών αποτυπωμάτων, διάγνωση μολυσματικών ασθενειών, κλωνοποίηση γονιδίων, δοκιμή πατρότητας, και υπολογισμός DNA .[2]
Η PCR ανακαλήφθηκε από Kary Mullis. Τον χρόνο που σκέφτηκε την PCR το 1983, ο Mullis εργαζόταν στο Emeryville,της Καλιφόρνιας για την Cetus, μιάς από τις πρώτες επιχειρήσεις βιοτεχνολογίας. Εκεί, χρεώθηκε με την παραγωγή των κοντών αλυσίδων του DNA για άλλους επιστήμονες. Ο Mullis έχει γράψει ότι συνέλαβε την PCR ταξιδεύοντας κατά μήκος της εθνικής οδού των ακτών του Ειρηνικού μια νύχτα στο αυτοκίνητό του [3]. Έπαιζε στο μυαλό του με έναν νέο τρόπο αλλαγών (mutations) στο DNA όταν συνειδητοποίησε ότι είχε αντ' αυτού ανακαλύψει μια μέθοδος για την ενίσχυση οποιωνδήποτε DNA περιοχών. Ο Mullis έχει πει ότι προτού να τελειώσει το ταξίδι του, ήταν ήδη σίγουρος για το Βραβείο Νόμπελ. Μοιράστηκε Βραβείο Νόμπελ στη χημεία με τον Michael Smith το 1993.[2]
Καθώς ο Mullis έγραψει στο Scientific American: " Αρχίζοντας με ένα απλό μόριο από το γενετικό υλικό του DNA, η PCR μπορέστε να παραγάγει 100 δισεκατομμύρια παρόμοια μόρια σε ένα απόγευμα. Η αντίδραση είναι εύκολο να εκτελεσθεί. Απαιτεί λιγότερος από ένα δοκιμαστικό σωλήνα, μερικά απλά αντιδραστήρια, και μια πηγή θερμότητας."[2]

Εικόνα 1: Ένα θερμικό cycler για PCR [4]
Η PCR χρησιμοποιείται για να ενισχύσει τις συγκεκριμένες περιοχές από ένα DNA κλώνο. Αυτό μπορεί να είναι ένα απλό γονίδιο, ακριβώς ένα μέρος ενός γονιδίου, ή μιά μη - κωδικοποιούσα ακολουθία. Η PCR χαρακτηριστικά ενισχύει μόνο μικρά DNA τεμάχια, συνήθως μέχρι 10 kilo base pairs (kb). Ορισμένες μέθοδοι μπορούν να αντιγράψουν τεμάχια πάνω από 25 kb σε μήκος που είναι ακόμα πολύ λιγότερο από το χρωμοσωμικό DNA από το ευκαριωτικό κύτταρο - παραδείγματος χάριν, ένα ανθρώπινο κύτταρο περιέχει περίπου τρία δισεκατομμύρια ζευγάρια βάσεων (3 Gbp). Η PCR, όπως αυτήν την περίοδο εφαρμόζεται, απαιτεί διάφορα βασικά συστατικά [5].
Αυτά τα συστατικά είναι:
· DNA πρότυπο, που περιέχει το τμήμα του DNA που ενισχύεται.
· Δύο εκκινητές, (βιοτινυλιωμένοι) έναν forward και έναν reverse, οι όποιοι καθορίζουν την αρχή και το τέλος της περιοχής που θα ενισχυθεί.
· Taq πολυμεράση (ή άλλη θερμοανθεκτική πολυμεράση), η DNA πολυμεράση που αντγιράφει την περιοχή που θα ενισχυθεί.
· Τριφωσφορικά δεοξυνουκλεοτίδια, (dNTPs) από τα οποία η DNA πολυμεράση χτίζει το νέο DNA (2Mm).
· Ρυθμιστικό διάλυμα, που παρέχει ένα κατάλληλο χημικό περιβάλλον για την DNA πολυμεράση [(x10) 100mM Tris-HCl, Ph 8.3 – 500mM KCl – 15mM MgCl (προαιρετικά)].
· Mg+2 κατιόν μαγνησίου (σε μορφή MgCl2
· Μονοσθενή κατιόντα καλίου
· Αποστειρωμένο απιονισμένο νερό.
· Πάγο.
· Αγαρόζη.
· 6X διάλυμα φόρτωσης δειγμάτων.
· Διάλυμα βρωμιούχου εθιδίου (10mg/ml). [6]
Η PCR διαδικασία πραγματοποιείται στο θερμικό cycler.(Εικόνα 1) Αυτός είναι μια μηχανή που θερμαίνει και ψύχει τους σωλήνες αντίδρασης μέσα στην ακριβή θερμοκρασία που απαιτείται για κάθε βήμα της αντίδρασης. Το καπάκι του θερμικού cycler θερμαίνεται για να αποτρέψει συμπύκνωση στο εσωτερικό από τα καλύμματα των δοκιμαστικών σωλήνων. Εναλλακτικά, ένα στρώμα του λαδιού μπορεί να τοποθετηθεί στο μίγμα αντίδρασης για να αποτρέψει την εξάτμιση. Χαρακτηριστική σειρά όγκων αντίδρασης από 15-100 μl. Αυτές οι μηχανές κόστος περισσότερο από $2,500 USD, από το 2004.[2]
Το τμήμα του DNA που θα ενισχυθεί καθορίζεται με την επιλογή των εκκινητών (primers). Οι εκκινητές είναι κοντοί, τεχνητοί DNA κλώνοι — συχνά με μήκος όχι περισσότερο από 50 και συνήθως μόνο 18 έως 25 ζεύγη βάσεων— που είναι συμπληρωματικοί στην αρχή ή το τέλος του τμήματος του DNA που θα ενισχυθεί. Αυτοί ανασυνδιάζονται με πρόσδεση στο DNA πρότυπο στα αρχικά και τελικά σημεία, όπου η DNA πολυμεράση δεσμεύεται και αρχίζει τη σύνθεση του νέου κλώνου του DNA.[2]
Η επιλογή του μήκους από τους εκκινητές και τη θερμοκρασία τήξης (Tm) εξαρτάται από έναν αριθμό από εκτιμήσεις. Η θερμοκρασία τήξης του εκκινητή - για να μην συγχεθεί με τη θερμοκρασία τήξης του προτύπου DNA - ορίζεται ως η θερμοκρασία κατά την οποία οι μισές θέσεις σύνδεσης των εκκινητών είναι κατειλημμένες. Οι εκκινητές που είναι πάρα πολύ κοντοί συνδέονται σε διάφορες θέσεις στο μεγάλο DNA πρότυπο, και θα οδηγήσουν σε μη - συγκεκριμένα αντίγραφα. Αφ' ετέρου, το μήκος του εκκινητή περιορίζεται από τη μέγιστη θερμοκρασία που θα εφαρμοστεί προκειμένου να το λειώσει, καθώς η θερμοκρασία τήξης αυξάνει με το μήκος του εκκινητή. Θερμοκρασίες τήξης που είναι πάρα πολύ υψηλές, π.χ., άνω των
Μερικές φορές οι εκφυλισμένοι εκκινητές χρησιμοποιούνται. Αυτά είναι πραγματικά μίγματα από παρόμοιους, αλλά όχι ίδιους, εκκινητές. Μπορούν να είναι κατάλληλοι εάν το ίδιο γονίδιο πρόκειται να ενισχυθεί από διαφορετικούς οργανισμούς, σαν γονίδια οι ίδιοι είναι πιθανώς παρόμοιοι αλλά όχι ίδιοι. Η άλλη χρήση για εκφυλισμένους εκκινητές είναι όταν των εκκινητών η σχεδίαση είναι βασισμένη στην πρωτεϊνική ακολουθία. Όπως ξέρουμε αρκετά διαφορετικά κωδικόνια μπορούν να κωδικοποιήσουν για έναν αμινοξύ, και είναι συχνά δύσκολο να συμπεράνουμε ποιό κωδικόνιο χρησιμοποιείται σε κάθε περίπτωση. Επομένως η αντιστοιχία της ακολουθίας του εκκινητή για το αμινοξύ ισολευκίνη μπορεί να είναι "ATH", όπου A κλώνος για την αδενίνη, T για την θυμίνη, και H για την αδενίνη, θυμίνη, ή κυτοσίνη. Χρήση εκφυλισμένων εκκινητών μπορεί πολύ να μειώσει την ελειδίκευση της PCR ενίσχυσης. Αυτό το πρόβλημα μπορεί να λυθεί με τη χρησιμοποίηση touchdown pcr [2]
Οι προαναφερθείσες εκτιμήσεις καθιστούν τη σχεδίαση του εκκινητή μια πολύ επίπονη διαδικασία, με την οποία παράγεται ένα προϊόν:
· Η GC-περιεκτικότητα πρέπει να είναι μεταξύ 40-60%.
· Η υπολογισμένη Tm και για τους δύο χρησιμοποιούμενους εκκινητές στην αντίδραση δεν πρέπει να διαφέρει περισσότερο από
· Θερμοκρασία αναδιάταξης συνήθως είναι
· Πρέπει να αποφευχθεί εσωτερική αυτοσυμπληρωματική φουρκέτα από >4 και από >8 διμερή. Η σχεδίαση του 3'άκρου του εκκινητή είναι κρίσιμο στην επιτυχία της PCR από εκκινητή που επιμηκύνεται από το 3' άκρο. Το 3΄ άκρο δεν πρέπει να είναι συμπληρωματικό με περισσότερες από 3-4 βάσεις σε οποιαδήποτε περιοχή του άλλου εκκινητή (ή ακόμα και του ίδιου εκκινητή) που χρησιμοποιείται στην αντίδραση και πρέπει να παρέχει σωστό ταίριασμα βάσεων στο πρότυπο.
Υπάρχουν προγράμματα υπολογιστών για να βοηθήσουν στον σχεδιασμό των εκκινητών.[2]
Ένα από αυτά τα προγράμματα (Εικόνα 2,3)είναι και το GeneACT.[7]

Εικόνα 2 PCR Γεννήτρια εκκινητών[7]

Εικόνα 3 Πιθανό αποτέλεσμα περιοχών συνδέσεων [7]
1. Exo- Pfu DNA Polymerase |
2. Herculase Enhanced DNA Polymerase |
3. Pfu DNA Polymerases |
4. PfuTurbo DNA Polymerase |
5. SureStart Taq DNA Polymerase |
6. Taq DNA Polymerase |
7. Taq2000™ DNA Polymerase |
8. TaqPlus Long PCR System |
9. TaqPlus Precision PCR System |
10. YieldAce DNA Polymerase |
· Γενετικά κατασκευασμένη μετάλλαξη από κλωνοποιημένη Pfu DNA πολυμεράση
· Μη ανιχνεύσιμη 3' πρός 5' διόρθωση δραστηριότητας εξωνουκλεάσης
· Ενσωματώνει [35S]dATP 10 φορές περισσότερο αποτελεσματικά από την Taq DNA πολυμεράση
· Αποτελεσματικά ενισχύει προβληματικούς, σύνθετους ή σε GC-πλούσιους στόχους
· Περιέχει τον ArchaeMaxx παράγοντα ενίσχυσης
· Υψηλότερη πιστότητα από την Taq DNA πολυμεράση και τα Taq- βασισμένα μίγματα
· Ιδανική για υψηλής πιστότητας ενίσχυση
· 3'-5' δραστηριότητα εξωνουκλεάσης παρέχει ένα χαμηλό ποσοστό λάθους
· Ένας από τις πιό γνωστές θερμοάντοχες DNA πολυμεράσες
· Έλλειψη επέκτασης βάσεων δραστηριότητα σημαίνει καμμία ανεπιθύμητη
· Βέλτιστος για blunt-end PCR κλωνοποίηση
· Βέλτιστη θερμοκρασία κοντά στους
· 95% ενεργή μετά από τη μίας ώρας επώαση σε
· Ειδική «έκδοση» από την κλωνοποιημένη Pfu DNA πολυμεράση και τον αποκλειστικός ArchaeMaxx πολυμεράσης παράγοντας ενίσχυσης 1
· Ίδια υψηλή πιστότητα όπως η Pfu DNA πολυμεράση με την πιό γερή απόδοση
· Ενισχύει PCR παραγωγές και ρυθμοαπόδοση προϊόντων
· Ενισχύει μακρύτερους στόχους από άλλα υψηλής πιστότητας ένζυμα
· Απαιτεί τους πιό σύντομους χρόνους επέκτασης, λιγότερους PCR κύκλους και λιγότερο DNA
· Ιδανικό για την ενίσχυση σύνθετων γενωμικών στόχων DNA μέχρι 19 kb και πλασμιδιακοί στόχοι μέχρι 15 kb
· Hot-start έκδοση διαθέσιμη για την εξειδίκευση της ενίσχυσης
· Hot start διατύπωση της Taq DNA πολυμεράσης
· Μεγαλύτερη PCR εξειδίκευση
· Μειωμένο μη συγκεκριμένο υπόβαθρο
· Βελτιωμένος προϊόν παραγωγής
· Ευπροσάρμοστος - μπορεί να χρησιμοποιηθεί για αργή ή γρήγορη ενεργοποίηση
· Αξιόπιστο setup σε θερμοκρασία δωματίου -
· Χρησιμοποιείται σε συνήθη PCR πρωτόκολλα
· Ιδιαίτερα καθαρισμένη φυσική Taq DNA πολυμεράση
· Κατάλληλη όταν η πιστότητα δεν είναι κρίσιμη
· Απομονωμένος από τα θερμοφιλικά ευβακτήρια Thermus aquaticus strain yT-1
· Αντιστέκεται σε θερμοκρασίες μέχρι
|
Εικόνα 4 Οι θέσεις DARNDEST: Οι επιστήμονες απομόνωσαν θερμοάντοχη DNA πολυμεράση Taq, ένα ένζυμο που οδηγεί την PCR, από το Thermus aquaticus Yellowstone type-1,(Εικόνα 4) έναν κάτοικο του θερμοπίδακα όπως αυτό στο Yellowstone National Park. [8] |

· Εξαιρετικά καθαρά κλωνοποιημένη Taq DNA πολυμεράση
· Ουσιαστικά αποβάλλει υπόβαθρου ψευδή ευρήματα
· Ελαχιστοποιεί το επίχρισμα στην long PCR
· Ιδανική για extreme PCR καταστάσεις
· Περισσότερο θερμοάντοχη από την Taq DNA πολυμεράση
TaqPlus Long PCR Σύστημα
· Ενισχύει τα πρότυπα μέχρι 35 kb
· Βελτιστοποιημένο μίγμα από Pfu και Taq2000 DNA πολυμεράσες
· Πιό αξιόπιστη από την Taq DNA πολυμεράση
· Χρόνοι επέκτασης τόσο σύντομοι όπως 30 seconds
TaqPlus Ακρίβειας PCR Σύστημα
· 2.7 φορές ακριβέστερη από την Taq DNA πολυμεράση
· Βελτιστοποιημένο μίγμα από Pfu και Taq2000 DNA πολυμεράσες
· Ενισχύει πρότυπα μέχρι 15 kb
· Κατά μέσος όρο 3- επίπεδα μεγαλύτερες παραγωγές από την Taq DNA πολυμεράση
· Ιδανική για την ενίσχυση σε βακτηριακή καλλιέργεια
· Υψηλότερη παραγωγή από την Taq DNA πολυμεράση από ποικίλα πρότυπα
· YieldAce Hotstart DNA πολυμεράση παρέχει υψηλή εξειδίκευση με setup σε θερμοκρασίες δωματίου. [9]
Η PCR διαδικασία συνήθως αποτελείται από μια σειρά από είκοσι έως τριάντα - πέντε κύκλους. Κάθε κύκλος αποτελείται από τρία βήματα (Εικόνα 5 ).
· Το δίκλωνο DNA πρέπει να θερμανθεί σε 94-
· Μετά από το χωρισμό των DNA κλώνων, η θερμοκρασία μειώνεται, έτσι οι εκκινητές μπορούν να συνδεθούν με το μονόκλωνο DNA. Αυτό το βήμα καλείται υβριδοποίηση. Η θερμοκρασία αυτού του σταδίου εξαρτάται από τους εκκινητές και είναι συνήθως
· Τέλος, η DNA πολυμεράση πρέπει να αντιγράψει τους DNA κλώνους. Αρχίζει υβριδοποιεί τον εκκινητήρα και να δουλεύει κατά μήκος του DNA κλώνου. Αυτό το βήμα καλείται επιμήκυνση. Η θερμοκρασία επιμήκυνσης εξαρτάται από την DNA πολυμεράση. Πολυμεράση Taq επιμηκύνει βέλτιστα σε μια θερμοκρασία 72 βαθμών Κελσίου. Ο χρόνος για αυτό το βήμα εξαρτάται και αυτός από την DNA πολυμεράση το ίδιο και για το μήκος των τμημάτων του DNA για θα ενισχυθεί. Εμπειρικά, αυτό το βήμα παίρνει 1 λεπτό ανά χίλια ζεύγη βάσεων. Ένα τελικό βήμα επιμήκυνσης χρησιμοποιείται συχνά μετά από τον τελευταίο κύκλο για να εξασφαλίσει ότι οποιαδήποτε παραμονή απλόυ κλώνου DNA αντιγράφεται εντελώς. Αυτό διαφέρει από όλα τα άλλα βήματα της επιμήκυνσης μόνο δεδομένου ότι είναι χαρακτηριστικά μακρύτερο 10-15 λεπτά. Αυτό το τελευταίο βήμα συστήνεται ιδιαίτερα εάν το pcr προϊόν είναι συνδεδεμένο με T φορέα με χρήση TA- κλωνοποίησης.

Εικόνα 5 : σχέδιο του pcr κύκλου. (1) Μετουσίωση σε 94-
Οι χρόνοι και οι θερμοκρασίες που δίνονται σε αυτό το παράδειγμα λαμβάνεται από ένα pcr πρόγραμμα που χρησιμοποιήθηκε επιτυχώς σε ένα 250 bp τμήμα από C- τελικό άκρο του αύξητικού παράγοντα όπως στην ινσουλίνη (IGF).

Εικόνα 6 "απεικόνιση " DNA μέσω της ηλεκτροφόρησης πηκτωμάτων αγαρόζης του προϊόντος αλυσιδωτής αντίδρασης πολυμεράσης PCR [11]
Το μίγμα της αντίδρασης αποτελείται από:
· µl DNA template (100 ng/µl)
· 2.5 µl of primer, 1.25 µl per primer (100 ng/µl)
· µl Pfu-Polymerase
· µl nucleotides
· 5.0 µl buffer solution
· 89.5 µl water
Ένας 200 µl σωλήνας αντίδρασης που περιέχει 100 µl μίγμα εισάγεται στον thermocycler.
Η PCR διαδικασία αποτελείται από τα ακόλουθα βήματα συνοπτικά:
· Έναρξη. Το μίγμα θερμαίνεται στους
· Τήξη, όπου θερμαίνεται σε
· Υβριδοποίηση (επαναδιάταξη με τη θέρμανση σε
· Επιμήκυνση με θέρμανση σε
Οι βάσεις (συμπληρωματικές στο πρότυπο) συνδέονται με τον εκκινητή στο
· Τα βήματα 2-4 επαναλαμβάνονται 25 φορές, αλλά με καλούς εκκινητές και φρέσκια πολυμεράση, 15 to 20 κύκλοι είναι ικανοποιητικοί.
· Το μίγμα κρατιέται σε
Το pcr προϊόν μπορεί να προσδιοριστεί από το μέγεθός του με χρήση ηλεκτροφόρησης πηκτωμάτων αγαρόζης.(Εικόνα 6) Η ηλεκτροφόρηση πηκτωμάτων αγαρόζης είναι μια διαδικασία που αποτελείται από την έγχυση του DNA στο πήκτωμα αγαρόζης και εφαρμόζοντας μετά ένα ηλεκτρικό ρεύμα στο πήκτωμα. Κατά συνέπεια, οι μικρότεροι DNA κλώνοι κινούνται γρηγορότερα από τους μεγαλύτερους κλώνους μέσω του πηκτώματος προς τον θετικό πόλο. Το μέγεθος του pcr προϊόντος μπορεί να καθοριστεί με τη σύγκριση του με μιά DNA κλίμακα, που περιέχει DNA τμήματα μεγνωστά μήκη, επίσης μέσα στο πήκτωμα.[2]
Δεδομένου ότι η pcr είναι πολύ ευαίσθητη, επαρκή μέτρα πρέπει να ληφθούν για να αποφευχθεί η μόλυνση από άλλο παρόν DNA στο περιβάλλον των εργαστηρίων (βακτηρίδια, ιοί, προσωπικό εργαστηρίων δέρμα κλπ.). Κατά συνέπεια η προετοιμασία των δειγμάτων DNA, το μίγμα του συνόλου αντίδρασης και η pcr διαδικασία, επιπλέον στην επόμενη αντίδραση της ανάλυση των προϊόντων, πρέπει να εκτελελούνται σε χωριστές περιοχές. Για την προετοιμασία από το μίγμα αντίδρασης, συστήνεται ένα γραφείο ελασματικής ροή με UV λαμπτήρα. Τα φρέσκα γάντια πρέπει να χρησιμοποιηούνται για κάθε ένα PCR βήμα επίσης όπως και η μετακίνηση των pipettes με τα φίλτρα αερολύματος. Τα αντιδραστήρια για την PCR πρέπει να προετοιμάζονται χωριστά και να χρησιμοποιούνται απλώς για αυτόν το λόγο. Αντιπροσωπευτικά δείγματα πρέπει να αποθηκεύονται χωριστά από άλλα δείγματα DNA. Μια αντίδραση ελέγχου (εσωτερικός έλεγχος), που εξαιρεί πρότυπο DNA, πρέπει πάντα να εκτελείται, για να επιβεβαιώνει την απουσία από τη μόλυνση ή σχηματισμός πολυμερών εκκινητών.
Βελτιστοποίηση ακινητοποίησης ολιγονουκλεοτιδίων ανιχνευτών.
Υπόστρωµα - Υψηλή οµοιογένεια - Χαµηλός αυτοφθορισµός.
Ολιγονουκλεοτίδιο ανίχνευσης.
Καθαρότητα→Ευαισθησία
Μήκος
Χηµεία σύνδεσης → Σταθερότητα κατά τα επόµενα στάδια υβριδισµού και έκπλυσης (θερµοκρασίες
Βελτιστοποίηση υβριδισµού ολιγονουκλεοτιδίων στόχων.
Επαρκής ενσωµάτωση µορίων σηµατοδότησης (ιχνηθετών) κατά την PCR.
Σύσταση του διαλύµατος υβριδισµού: ιονική ισχύς, αποδιατακτικοί παράγοντες, κλπ.
Θερµοκρασία υβριδισµούTm : θερµοκρασία στην οποία τα πλήρως συµπληρωµατικά ολιγονουκλεοτίδια βρίσκονται συµπλεγµένα κατά 50%
Td : θερµοκρασία σε µια συγκεκριµένη συγκέντρωση άλατος και ολιγονουκλεοτιδίων στην οποία το 50% ενός ολιγονουκλεοτιδίου είναι συµπλεγµένο µε το πλήρως συµπληρωµατικό του Td=
Δευτεροταγείς δομές DNA, προκαλούμενες από το ζευγάρωμα των βάσεων του ίδιου κλώνου του μορίου, μπορούν να προκαλέσουν το δίπλωμα ή ακόμα και το δέσιμο των προτύπων του DNA ή των εκκινητών, οδηγώντας σε μειωμένη παραγωγή ή συνολική αποτυχία της αντίδρασης. Φουρκέτοειδές, άμεσο δίπλωμα του DNA προκαλούμενο από τρέξιμο των συμπληρωματικών βάσεων ή μια αντιστροφή, είναι τα πιό κοινά προβλήματα αυτού του είδους.
Χαρακτηριστικά, αυτό απαιτεί την επιλογή διαφορετικών εκκινητών. Δευτεροταγείς δομές στο πρότυπο DNA δεν είναι τόσο σημαντικές όσο εκείνες των εκκινητών, καθώς η DNA πολυμεράση "ισοπεδώνει" τις περισσότερες δευτεροταγείς δομές εκτός και αν είναι ιδιαίτερα γερές.
Εντούτοις, εάν η χρήση των ελαττωματικών εκκινητών είναι απαραίτητη, όπως συμβαίνει συχνά στην περίπτωση που πρέπει να γίνει και κλωνοποίηση, το πρόβλημα μπορεί να βελτιωθεί κάπως μέσω της DMSO ή της γλυκερίνης. Αυτές οι χημικές ουσίες μπορούν να προστεθούν στην pcr mastermix για να διαπάσουν τις δευτεροταγείς δομές.[2]
Η Taq πολυμεράση στερείται 3' σε 5' εξωνουκλεασική δραστηριότητα. Αυτό καθιστά αδύνατο το να ελένχει τη βάση που έχει παρεμβάλει και να την αφαιρεί εάν δεν είναι η σωστή, μια διαδικασία κοινή στους ανώτερους οργανισμούς. Αυτό οδηγεί στη συνέχεια σε ένα υψηλό ποσοστό λάθους περίπου 1 στις 10,000 βάσεις, έτσι, εάν ένα λάθος εμφανίζεται νωρίς, μπορεί να αλλάξει τα μεγάλα ποσοστά του τελικού προϊόντος.
Άλλες πολυμεράσεις είναι διαθέσιμες για την ακρίβεια σε ζωτικής σημασίας χρήσεις όπως η ενίσχυση για sequencing. Παραδείγματα πολυμεράσεων με 3' σε 5' εξωνουκλεασική δραστηριότητα περιλαμβάνουν: KOD DNA πολυμεράση, μια ανασυνδυαζόμενη μορφή του Thermococcus kodakaraensis KOD1. Vent, που εξάγεται από το Thermococcus litoralis; Pfu DNA πολυμεράση, που εξάγεται από το Pyrococcus furiosus; και Pwo, που εξάγεται από το Pyrococcus woesii.[2]
Η PCR δουλεύει εύκολα με DNA με μήκος δύο με τρεις χιλιάδες ζεύγη βάσεων, αλλά αυτό το μήκος η πολυμεράση τείνει να το μειώσει, και ο χαρακτηριστικός κύκλος θέρμανσης δεν αφήνει αρκετό χρόνο για να ολοκληρωθεί ο πολυμερισμός. Είναι δυνατό να ενισχυθούν μεγαλύτερα κομμάτια μέχρι 50,000 ζέυγη βάσεων με έναν πιό αργό κύκλο θέρμανσης και ειδικές πολυμεράσες. Αυτές οι ειδικές πολυμεράσες είναι συχνά πολυμεράσες λιωμένες με παρατετταμένη - ενίσχυση DNA- σε σύνδεση με πρωτεϊνη, κάνοντας αυτές κυριολεκτικά "κολλητές" στο περισσότερο DNA [13][14].
Άλλες πολύτιμες ιδιότητες του πρωτοτύπου των χειμερικών πολυμερασών TopoTaq και PfuC2 περιλαμβάνουν ενισχυμένες θερμοανθεκτικές, εξειδικευμένες και με αντίσταση στους μολυσματικούς παράγοντες και στους ανασταλτικούς παράγοντες [15][16]. Κατασκευάστηκαν με χρήση μοναδικών Helix-hairpin-Helix (HhH) DNA δεσμευτικών περιοχών από τοποϊσομεράσες V [17] από υπερθερμόφιλα Methanopyrus kandleri. Χειμερικές πολυμεράσες υπερνικούν πολλούς περιορισμούς από τα εγγενή ένζυμα και χρησιμοποιούνται σε άμεση PCR ενίσχυση από κυτταροκαλλιέργειες και ακόμη και δείγματα τροφίμων κατά συνέπεια με εφαρμογή μιάς επίπονης διαδικασίας απομόνωσης του DNA. Μια γερή δραστηριότητα μετατόπισης κλώνων από το υβρίδιο TopoTaq πολυμεράση βοηθά να λύσει τα προβλήματα της PCR με φουρκέτες και GC- πλούσια πρότυπα [18].
Η μη συγκεκριμένη σύνδεση των εκκινητών είναι πάντα μια δυνατότητα λόγω των διπλασιασμών ακολουθίας, μη ειδικών συνδέσεων και μερικών συνδέσεων των εκκινητών, αφήνοντας τα
Ένας νέος τρόπος να διατηρηθούν θερμόφιλα ένζυμα απολύτως ανενεργά στη χαμηλή θερμοκρασία βρέθηκε κατά τη διάρκεια των δομικών μελετών ενός υπερθερμόφιλου ενζύμου συνδεόμενου με το DNA [19]. Ειδικά κατασκευασμένη TopoTaq πολυμεράση ενεργοποιείται αμέσως στην υψηλή θερμοκρασία και υπερνικά τους περιορισμούς του συμβατικό "hot-start" ενζύμου που πρέπει να περιμένει ένα αντίσωμα για να αποσπάσει ενώ ο DNA στόχος αποικοδομείται σε 95˚C. Επιπλέον, η δραστηριότητά του εμποδίζεται με την ολοκλήρωση της PCR στη χαμηλή θερμοκρασία.
Άλλες μέθοδοι για να αυξηθεί η ειδικότητα είναι η Nested PCR και η Touchdown PCR.
Πρακτικές τροποποιήσεις στη pcr τεχνική
Nested PCR
Η Nested PCR (Εικόνα 7,8) προορίζεται να μειώσει τις μολύνσεις στα προϊόντα λόγω της ενίσχυσης από απροσδόκητες περιοχές συνδέσεων εκκινητών. Δύο σύνολα εκκινητών χρησιμοποιούνται σε δύο διαδοχικά PCR τρεξίματα, το δεύτερο σύνολο είναι προορισμένο για να ενισχύσει έναν δεύτερο στόχο που το πρώτο τρέξιμο παρήγαγε. Αυτό είναι πολύ επιτυχές, αλλά απαιτεί περισσότερη λεπτομερή γνώση από τις σχετικές ακολουθίες.

Εικόνα 7 Διαγραμματική αναπαράσταση από το πρωτόκολλο nested-polymerase chain reaction (PCR) για την ενίσχυση μιας μερίδας του γονιδίου για το 16S ribosomal RNA. Σημειώστε ότι η πρώτη PCR ενισχύει περ. 1.8 kbp ακολουθία από το rRNA operon, και ότι η δεύτερη (nested) PCR ενισχύει περ. 1.2 kbp ακολουθία από το γονίδιο για το 16S rRNA. [20]

Εικόνα 8 Nested PCR [21]

Εικόνα 9 Αρχές της ISSR PCR [22]
Διά - απλή ακουκουθία επαναλήψεων αξιοποιείται από την απλή ακολουθία επαναλήψεων (SSR) ή τους μικροδορυφόρους που είναι άφθονοι σε όλα τα ευκαριωτικά γονιδιώματα. Οι SSR είναι κοντές, διαδοχικές επαναλήψεις που είναι χρήσιμοι ως γενετικοί δείκτες. Εντούτοις, η χρήση των SSR απαιτεί γνώση της ακολουθίας των περιοχών του πλαισίου των διαδοχικών επαναλήψεων. Η ISSR αντίθετα, δεν απαιτεί καθόλου προγενέστερη γνώση από την ακολουθία γονιδιώματος. Η ISSR χρησιμοποιεί εκκινητές που αναχωρούν από το 5' ή 3' άκρο της επαναλαμβανόμενης περιοχής και επεκτείνονται στην περιοχή πλαισίωσης. Αυτή η τεχνική κατόπιν επιτρέπει την ενίσχυση των γενωμικών τμημάτων μεταξύ αντιστρόφως προσανατολισμένων επαναλήψεων (ISSRs). Γενικά μια σειρά απλών εκκινητών χρησιμοποιείται για να παραγάγει τη σειρά τμημάτων που είναι που χωρίζονται ανάλογα με το μέγεθος είτε σε ένα πήκτωμα αγαρόζης είτε πολυακρυλαμιδίου.
Αυτή η τεχνική είναι γρήγορη και μπορεί να διαφοροποιηθεί μεταξύ πολύ σχετικών ατόμων. Πλεονεκτήματα από αυτήν την τεχνική αποτελούν οι πολλαπλάσιες θέσεις πολυμορφισμού, υψηλή ρυθμοαπόδοση και το χαμηλότερο κόστος. Μειονέκτημα αποτελεί το γεγονός ότι αυτοί είναι οι κυρίαρχοι δείκτες. [23]
FISSR-PCR
Με την τεχνική αυτή (FISSR-PCR) τα informativeness, που μετρήθηκαν σαν τον αριθμό των ανιχνεύσιμων τμημάτων που ενισχύεται, ήταν δύο φορές υψηλότερα και η ποσότητα από τα απαραίτητα πρότυπα DNA ήταν δύο φορές χαμηλότερα από την απλή ISSR-PCR. Έχουμε καλέσει αυτήν την μέθοδο σαν FISSR-PCR (Εικόνα 10) και είναι μεγάλη η χρησιμότητά της στην παραγωγή μεγάλου αριθμού ειδών και ποικίλων ειδικών δεικτών στα φυτά, στα έντομα, στα παράσιτα από τα έντομα και σε ανθρώπινους και διάφορους άλλους μολυσματικούς οργανισμούς.Αποδεικνύεται ότι οι FISSR δεικτες κληρονομούνται και διαχωρίζονται σαν Μεντελικοί χαρακτήρες. Η FISSR-PCR ανάλυση δείκτη θα μπορούσε να είναι μια μέθοδος επιλογής για τη διαλογή μεγάλης κλίμακας από ποικιλίες και μεγάλης έκτασης γενοτύπηση στη χαρτογράφηση των γονιδιωμάτων όπου οι μικροδορυφορικές πληροφορίες είναι παραγγειακές ή απούσες.[24]

Εικόνα 10 FISSR-PCR DNA σχέδιο finerprinting για Casurina equisetifollia κλώνους [25]

Εικόνα 11 . Διάγραμμα ροής αυτοματοποιημένης LM-PCR. (A) Αυτοματοποιημένη AmpliGrease hot-start LM-PCR είναι μια τροποποίηση manual πρωτόκολλου για τη χρήση της ζελατίνας πετρελαίου σαν εμπόδιο που χωρίζει δύο υδάτινα συστατικά μέχρι heated. Τα συστατικά είναι αριθμημένα με τάξη σε σωλήνες με χρήση χειρισμού ενός ρομπότ υγρού. Το πρώτο συστατικό είναι ένας περιορισμός προμίξεων PCR ρυθμιστικών διαλυμάτων, δεοξυνουκλεοτιδίων, ολιγονουκλεοτιδίων, εκκινητών, και πρότυπου, το οποίο μπορεί να προστεθεί χωριστά. Το ορυκτέλαιο εισάγεται με σιφώνιο στο σωλήνα σε δύο καταστάσεις, πρώτα σαν 7.5 ![]() l συστατικό για να προστατεύσει την κατώτατη πρόμιξη από το ράντισμα και για να απορροφήσει τη θερμότητα από τη λειωμένη ζελατίνα πετρελαίου, και έπειτα ως μεγαλύτερο υποπολλαπλάσιο για να επιτρέψει στη ζελατίνα πετρελαίου να διαμορφώσει μια ενιαία φάση στη θέρμανση. Τέλος, ένα δεύτερο υδάτινο υποπολλαπλάσιο εισάγεται με σιφώνιο στο ορυκτέλαιο που περιέχει την πολυμεράση. Εξίσωση των συγκεντρώσεων του απομονωτή και του Mg2+ και στις δύο υδάτινες φάσεις έγινε για να βελτιώσει την αξιοπιστία. (B) Η LM-PCR αυτοματοποιήθηκε με την αραίωση των προϊόντων από κάθε κατάσταση στην επόμενη χρησιμοποίηση AmpliGrease σε τρείς καταστάσεις όπως υποδεικνύεται από τα μπλε βέλη. (C) Παράδειγμα της ακολουθίας ανθρώπου TP53 εξωνίου-5 κατά μήκος του ανωτέρου κλώνου. Οι διάδρομοι (από το αριστερό στα δεξιά) είναι G, G+A, C+T, και C προϊόντα από Maxam−Gilbert25 αλληλούχισης αντιδράσεις διενεργηθείσες με χρήση HeLa κυττάρου DNA. Η ακολουθία που επιδεικνύεται βρίσκεται 137−251 bp από την ακολουθία των εκκινητών (P53R7028).[26]
l συστατικό για να προστατεύσει την κατώτατη πρόμιξη από το ράντισμα και για να απορροφήσει τη θερμότητα από τη λειωμένη ζελατίνα πετρελαίου, και έπειτα ως μεγαλύτερο υποπολλαπλάσιο για να επιτρέψει στη ζελατίνα πετρελαίου να διαμορφώσει μια ενιαία φάση στη θέρμανση. Τέλος, ένα δεύτερο υδάτινο υποπολλαπλάσιο εισάγεται με σιφώνιο στο ορυκτέλαιο που περιέχει την πολυμεράση. Εξίσωση των συγκεντρώσεων του απομονωτή και του Mg2+ και στις δύο υδάτινες φάσεις έγινε για να βελτιώσει την αξιοπιστία. (B) Η LM-PCR αυτοματοποιήθηκε με την αραίωση των προϊόντων από κάθε κατάσταση στην επόμενη χρησιμοποίηση AmpliGrease σε τρείς καταστάσεις όπως υποδεικνύεται από τα μπλε βέλη. (C) Παράδειγμα της ακολουθίας ανθρώπου TP53 εξωνίου-5 κατά μήκος του ανωτέρου κλώνου. Οι διάδρομοι (από το αριστερό στα δεξιά) είναι G, G+A, C+T, και C προϊόντα από Maxam−Gilbert25 αλληλούχισης αντιδράσεις διενεργηθείσες με χρήση HeLa κυττάρου DNA. Η ακολουθία που επιδεικνύεται βρίσκεται 137−251 bp από την ακολουθία των εκκινητών (P53R7028).[26]
Inverse PCR
Η Inverse PCR είναι μια μέθοδος που χρησιμοποιείται για να επιτρέψει την PCR όταν μόνο μια εσωτερική ακολουθία είναι γνωστή. Αυτό είναι ιδιαίτερα χρήσιμο στον προσδιορισμό ακολουθιών πλαισίωσης σε ποικίλα γονιδιακά ένθετα. Αυτό περιλαμβάνει μια σειρά από πέψεις και αυτο- συνδέσεις πριν από την κοπή από μια ενδονουκλεάση, με συνέπεια γνωστές ακολουθίες στο τέλος καθεμιάς από τις άγνωστες ακολουθίες.
Inverse- ή I-PCR είναι ένας τρόπος για την απομόνωση άγνωστων ακολουθιών πλαισίωσης από μια γνωστή ακολουθία. Η διαδικασία εξηγείται καλύτερα με το παρακάτω σχέδιο.
|
Εικόνα 12 [27] |

Παράδειγμα: Για την απομόνωση ανοδικού T-DNA ακολουθιών πλαισίωσης από τις γραμμές που μετασχηματίζονται με pGKB5 προαγωγέα κατασκευάζονται παγίδες γονιδίων (Versailles T-DNA lines). [27]

Εικόνα 13 [27]
Vectorette-PCR
Vectorette ή V-PCR είναι ένας άλλος τρόπος για την απομόνωση άγνωστων ακολουθιών πλαισίωσης από μια γνωστή ακολουθία. Η διαδικασία χρησιμοποιείται συνήθως στο εργαστήριο και εξηγείται καλύτερα από το σχέδιο κατωτέρω.

Εικόνα 14 (a) DNA με μια περιοχή της γνωστής ακολουθίας και πλαισιωμένη από τις ενζυμικές περιοχές περιορισμού R. (b) Ενζυμική διάσπαση του DNA σε τμήματα. (c) Η μονάδα vectorette επιδένεται σε όλα τα τμήματα DNA που προέκειψαν από την διάσπαση. (d) Ένας εκκινητήα συμπληρωματικός σε ένα κλώνο από τη γνωστή ακολουθία επεκτείνεται κατά τη διάρκεια της σύνθεσης του πρώτου κλώνου στον πρώτο pcr κύκλο. Αυτό παράγει ένα προϊόν του οποίου το 3' άκρο πλήρως συμπληρωματικό τώρα του vectorette συνδέεται στον κλώνο των προτύπων. (e) και (f) Δεύτερη σύνθεση κλώνων στον δεύτερο pcr κύκλο μπορεί μόνο να εμφανιστεί όπου vectorette συμπληρωματικές ακολουθίες έχουν παραχθεί. Δηλαδή τα προϊόντα από τον πρώτο κύκλο παράγονται από γνωστή - ακολουθία εκκινητή. (g) Οι επόμενοι κύκλοι pcr ενισχύουν το DNA οριακά από την γνωστή-ακολουθία εκκινητή και τον vectorette εκκινητή. Η αλληλούχιση μπορεί να διενεργηθεί από την αρχικη vectorette μονάδα με sp (εκκινητή αλληλούχισης).[27]

Εικόνα 15 [27]
RT-PCR
Η RT-PCR (Αντίστροφης μεταγραφής PCR) (Εικόνα 16,17,18) είναι η μέθοδος που χρησιμοποιείται για να ενισχύσει, απομονώσει ή προσδιορίσει μια γνωστή ακολουθία από το ένα κύτταρο ή ιστό βιβλιοθήκη RNA. Ουσιαστικά κανονική PCR προηγείται της αντίστροφης μεταγραφής (για να μετατρέψει το RNA σε cDNA) που χρησιμοποιείται ευρέως στην χαρτογράφηση της γονιδιακής έκφρασης, για το πότε και που ορισμένα γονίδια εκφράζονται.

Εικόνα 16 [28]

Εικόνα 17 RT-PCR [21]

Εικόνα 18 [29]
Assembly PCR
Η Assembly PCR είναι η απολύτως τεχνητή σύνθεση από τα μακριά προϊόντα γονιδίων με την εκτέλεση PCR σε μια δεξαμενή με μεγάλα ολιγονουκλεοτίδια με μικρά επικαλυπτόμενα τμήματα. Τα ολιγονουκλεοτίδια εναλλάσσονται μεταξύ κωδικής και αντικωδικής κατεύθυνσης, και η επικάλυψη τμημάτων χρησιμεύει για να διατάξει τα τεμάχια της PCR έτσι ώστε να παράγουν επιλεκτικά το τελικό προϊόν τους.[30]
Πρωτόκολλο για assembly PCR αντίδραση
Το πρόγραμμα (Εικόνα 19) ήταν πειραματικά ελεγχμένο με χρήση ολιγοδεοξυνουκλεοτιδίων καθορισμένων από το πρόγραμμα για την σε δύο στάδια assembly PCR κατασκευή ενός DNA μορίου που πρόκειται να χρησιμοποιηθεί για να παραγάγει ένα μόριο RNA. Το επιθυμητό προϊόν RNA είναι ένα 191-νουκλεοτιδίων μοριοαποτελούμενο από
Επάνω στην παραλαβή, τα ολιγοδεοξυνουκλεοτίδια για το πρώτο βήμα της assembly PCR αραιώθηκαν σε 0.125 µg/µL με το διπλά απεσταγμένο νερό, ενώ τα ολιγοδεοξυνουκλεοτίδια για δεύτεροpcr το βήμα αραιώθηκαν σε 0.25 µg/µL. Για την πρώτη pcr αντίδραση, 4 µL από κάθε έναν oligo, 4 µL από
Για τη δεύτερη pcr αντίδραση, 1 µL από το ακατέργαστο μίγμα από την πρώτη pcr αντίδραση αναμίχθηκε με 4 µL από κάθε εκκινητά, 4 µL από
Τα pcr μίγματα αναλύθηκαν από την ηλεκτροφόρηση πηκτωμάτων αγαρόζης. Για κάθε αντίδραση ένα 6 µL δείγμα αναμίχθηκε με 2 µL από μπλε - πράσινη χρωστική ουσία. Το πήκτωμα χρωματίστηκε με βρωμίδιο του εθιδίου για 20 λεπτά, και παρατηρήθηκε κάτω από το UV φως. Όπως παρουσιάζεται από τις αναλύσεις πηκτωμάτων (Εικόνα 19), η πρώτη pcr αντίδραση παράγει μια διάχυτη ζώνη ή μια κηλίδα, ενώ το επιθυμητό πλήρους μήκους προϊόν προκείπτει από τη δεύτερη pcr αντίδραση. Αυτή η συμπεριφορά είναι σύμφωνη με τις προηγούμενες εκθέσεις της assembly PCR κατασκευής γονιδίων. Το προϊόν της δεύτερης pcr αντίδρασης κλωνοποιήθηκε μέσα σε pUC18 πλασμίδιο, και η ακρίβεια της ακολουθίας του ελέγχθηκε από DNA αλληλούχιση.[30]

Εικόνα 19.[30]
Asymmetric PCR-Late PCR
Asymmetric PCR (Εικόνα 20) χρησιμοποιείται για να ενισχύσει κατά προτίμηση έναν κλώνο από το αρχικό DNA περισσότερο από τον άλλο. Βρίσκει χρήση σε μερικούς τύπους αλληλούχισης και σε εξέταση υβριδοποίησης όπου έχουμε μόνο ένα από τους δύο συμπληρωματικούς κλώνους, είναι ιδανική. Η PCR πραγματοποιείται ώς συνήθως, αλλά με μια μεγάλη υπερβολή εκκινητών για τον επιλεγμένο κλώνο. Λόγω της αργής (αριθμητικής) ενίσχυσης αργότερα στην αντίδραση μετά από τον περιορισμό του εκκινητή που έχει καταναλωθεί, πρόσθετοι κύκλοι PCR απαιτούνται. Μια πρόσφατη τροποποίηση σε αυτήν την διαδικασία, γνωστή σαν Linear-After-The-Exponential-PCR (LATE-PCR), χρησιμοποιεί έναν περιορισμό εκκινητή με μια υψηλότερη θερμοκρασία τήξης (Tm) από αυτήν της υπερβολής του εκκινητή για να διατηρήσει την αποδοτικότητα της αντίδρασης όπως κατά τον περιορισμό του εκκινητή.[21]

Εικόνα 20 Asymmetric PCR [21]
Quantitative PCR
Q-PCR (Quantitative PCR) (Εικόνα 21) χρησιμοποιείται για να μετρήσει γρήγορα την ποσότητα από το pcr προϊόν (κατά προτίμηση σε πραγματικό χρόνο), κατά συνέπεια είναι μια έμμεση ποσοτική μέθοδος που μετράει τα αρχικά ποσά του DNA, cDNA ή RNA. Αυτή χρησιμοποιείται συνήθως με σκοπό τον καθορισμό εάν μια ακολουθία είναι παρούσα ή όχι, και εάν είναι παρούσα να μετρήσει τον αριθμός των αντιγράφων στο δείγμα. Υπάρχουν 3 κύριες μέθοδοι που ποικίλλουν ως προς τη δυσκολία και τις λεπτομέρειες.

Εικόνα 21 Plexor qPCR & qRT-PCR [31]
Quantitative real-time PCR
Quantitative real-time PCR (Εικόνα 22) είναι συχνά γνωστή σαν RT-PCR (Real Time PCR) και RQ-PCR. QRT-PCR ή RTQ-PCR είναι πιό κατάλληλες συντομογραφίες. Σαν RT-PCR μπορούμε επίσης να αναφέρουμε τη reverse transcription PCR, που ακόμα πιό μπερδεμένα, χρησιμοποιείται συχνά από κοινού σαν Q-PCR. Αυτή η μέθοδος χρησιμοποιεί φθορισμού χρωστικές ουσίες και δείκτες για να μετράνε το ποσό του ενισχυμένου προϊόντος σε πραγματικό χρόνο.

Εικόνα 22 [32]
Δύο κοινές βασισμένες στον φθορισμό μέθοδοι ανίχνευσης DNA χρησιμοποιούνται για την Real Time PCR : 1) άμεσο μαρκάρισμα του dsDNA με SYBR Green I, ή 2) υβριδοποίηση με TaqMan δείκτες.
Taqman PCR
Ο TaqMan δείκτης είναι το κύριο χαρακτηριστικό της real-time PCR ανάλυσης και αποτελείται από ένα όχι-με δυνατότητα επέκτασης συμπληρωματικό ολιγονουκλεοτίδιο στην ακολουθία στόχων που μαρκάρεται στο 5'- με χρωστική ουσία αναφοράς και στο 3' με μια χρωστική ουσία απόσβεσης. (Εικόνα 23) Η φθορισμού χρωστική ουσία 6-carboxy-fluorescein (FAM), γενικά χρησιμοποιείται σαν χρωστική αναφοράς και είναι ομοιοπολικά συνδεδεμένη στο

Εικόνα 23 [34]

Εικόνα 24 [34]

Εικόνα 25 [35]
Η SYBR Green (Εικόνα 24,25) είναι βασική μέθοδος ανίχνευσης που επιτρέπει μεμονωμένα έναν έλεγχο εξειδίκευσης των αντιδράσεων. Το Panel A δείχνει την πρώτη-παράγωγο της καμπύλης τήξης για τις αντιδράσεις που κάθε μια χαρακτηρίζει την έκφραση ενός διαφορετικού μέλους από την BMP οικογένεια γονιδίων. Σε κάθε περίπτωση, η καμπύλη περιέχει μόνο μια μέγιστη ένδειξη διότι η αντίδραση παράγει μόνο ένα προϊόν. Το Panel B επιβεβαιώνει τα αποτελέσματα των καμπυλών τήξης δείχνοντας ότι αυτές οι ίδιες αντιδράσεις επίσης παράγουν μόνο μια ζώνη του προβλεφθέντος μεγέθους από την ηλεκτροφόρηση πηκτωμάτων αγαρόζης. Το Panel C περιλαμβάνει ένα παράδειγμα set εκκινητών που ανίχνευσε μια άγνωστη προηγουμένως παραλλαγή συναρμογών ή εναλλακτικό αντίγραφο από ένα γονίδιο με την παραγωγή ενός δεύτερου ενισχυμένου προϊόντος. Κατά συνέπεια, δύο αιχμές εμφανίζονται στην πρώτη παράγωγο της καμπύλης τήξης και δύο ζώνες εμφανίζονται από την ηλεκτροφόρηση πηκτωμάτων αγαρόζης. [35]
Touchdown PCR
Touchdown PCR (Εικόνα 26) είναι μια παραλλαγή pcr που μειώνει μη ειδικό υβριδισμό εκκινητή με πιό βαθμιαίο χαμήλωμα της θερμοκρασίας επαναδιάταξης μεταξύ των κύκλων. Δεδομένου ότι οι υψηλότερες θερμοκρασίες δίνουν μεγαλύτερη εξειδίκευση για την σύνδεση του εκκινητή, οι εκκινητές υβριδίζουν πρώτοι καθώς η θερμοκρασία περνά μέσα από τη ζώνη με τη μέγιστη εξειδίκευση.

Εικόνα 26 Αντιπρωσοπευτική Touchdown RT-PCR σε Synaptoneurosomal [36]
Hot-start PCR
Hot-start PCR (Εικόνα 27) είναι μια τεχνική που μειώνει το μη συγκεκριμένο εμπύρευμα που εμφανίζεται κατά τη διάρκεια της προετοιμασίας από τα τμήματα της αντίδρασης. Η τεχνική μπορεί να εκτελεσθεί με το χέρι με απλή θέρμανση των τμημάτων της αντίδρασης εν συντομία στη θερμοκρασία τήξης πρίν προστεθεί η πολυμεράση. Εξειδικευμένα ενζυμικά συστήματα έχουν αναπτυχθεί που εμποδίζουν τη δραστηριότητα της πολυμεράσης στην περιβαλλοντική θερμοκρασία, είτε με τη σύνδεση αντισωμάτων ή με την παρουσία ομοιοπολικά συνδεδεμένων ανασταλτικών παραγόντων που χωρίζουν μόνο μετά από έναν βήμα ενεργοποίησης υψηλής θερμοκρασίας. Hot-start/cold-finish PCR επιτυγχάνεται με τις νέες υβριδικές πολυμεράσες που είναι ανενεργές στην περιβαλλοντική θερμοκρασία και ενεργοποιούνται αμέσως στη θερμοκρασία επιμήκυνσης.

Εικόνα 27 Takara Hot Start PCR [37]
Η Taq πολυμεράση παρουσιάζει κάποια δραστηριότητα ακόμη και στις θερμοκρασίες σημαντικά κάτω από το βέλτιστό της. Επομένως, ανεπιθύμητες δευτερογενείς επεκτάσεις μπορούν να εμφανιστούν κατά τη διάρκεια των αντιδράσεων που συγκεντρώνονται σε θερμοκρασία δωματίου με συνέπεια τα πλαστά προϊόντα και το υψηλό υπόβαθρο. Αυτές οι ανεπιθύμητες επεκτάσεις μπορούν να αποφευχθούν με τη συγκέντρωση των αντιδράσεων στον πάγο, ή με χρήση Hot Start DNA πολυμεράσης.
Takara's Ex Taq, Taq και
Colony PCR
Βακτηριακοί κλώνοι (E.coli) μπορούν να προβληθούν με τα σωστά προϊόντα σύνδεσης. Επιλεγμένες αποικίες επιλέγονται με μία αποστειρωμένη οδοντογλυφίδα από ένα πιάτο αγαρόζης και αναμειγνύονται στο κύριο μίγμα ή το αποστειρωμένο ύδωρ. Εκκινητές (και το κύριο μίγμα) προστίθενται. Το PCR πρωτόκολλο πρέπει να αρχίσει με έναν παραταθέντα χρόνο σε 95˚C όταν η τυποποιημένη πολυμεράση χρησιμοποιείται ή με βήμα σύντομης αποδιάταξης σε 100˚C και ειδική χειμερική DNA πολυμεράση [18].

Εικόνα 28 [38]
Η Colony Fast-Screen (PCR Screen) (Εικόνα 28) γρήγορα προετοιμάζει κλώνους για τη διαλογή με PCR σε περίπου 10 λεπτά χωρίς την ανάγκη για καλλιέργειες ή DNA καθαρισμούς.[38]

Εικόνα 29 [39]
Η RACE-PCR (Εικόνα 29) διευκολύνει την απομόνωση από ![]() επέκταση ακολουθίας (ακολουθία αγκύρων, συχνά >15 νουκλεοτίδια μήκος) που ενσωματώνεται στο cDNA αντίγραφο στο βήμα της αντίστροφης μεταγραφής. Ένας εσωτερικός κωδικός εκκινητής χρησιμοποιείται έπειτα για να παραγάγει έναν κοντό δεύτερο κλώνο που τελειώνει σε μια συμπληρωματική ακολουθία στην αρχική ακολουθία αγκύρων. Έκτοτε, η PCR αρχίζει χρησιμοποιώντας τον εσωτερικό κωδικό εκκινητή και μια ακολουθία αγκύρων εκκινητή. (B)
επέκταση ακολουθίας (ακολουθία αγκύρων, συχνά >15 νουκλεοτίδια μήκος) που ενσωματώνεται στο cDNA αντίγραφο στο βήμα της αντίστροφης μεταγραφής. Ένας εσωτερικός κωδικός εκκινητής χρησιμοποιείται έπειτα για να παραγάγει έναν κοντό δεύτερο κλώνο που τελειώνει σε μια συμπληρωματική ακολουθία στην αρχική ακολουθία αγκύρων. Έκτοτε, η PCR αρχίζει χρησιμοποιώντας τον εσωτερικό κωδικό εκκινητή και μια ακολουθία αγκύρων εκκινητή. (B)
Η χρήση των πολλαπλών, μοναδικών sets εκκινητών μέσα σε μια απλή PCR αντίδραση (Εικόνα 30) για να παραγάγει ενισχυμένα τμήματα με ποικίλα μεγέθη ειδικά για διαφορετικές ακολουθίες DNA. Με το να στοχεύσει στα πολλαπλά γονίδια αμέσως, πρόσθετες πληροφορίες μπορούμε να αποσπάσουμε από το τρέξιμο μιας απλής δοκιμής που ειδάλλως θα απαιτούσε αρκετές φορές τα αντιδραστήρια και τον χρόνο των τεχνικών για να εκτελέσουν. Οι θερμοκρασίες υβριδοποίησης για κάθε ένα από τα sets των εκκινητών πρέπει να βελτιστοποιηθούν για να δουλέψει σωστά μέσα σε μια απλή αντίδραση και τα μεγέθη των ενισχυμένων τμημάτων πρέπει να χωριστούν λόγω αρκετής διαφοράς στο τελικό μήκος ζευγαριών βάσεων για να διαμορφώσει τις ευδιάκριτες ζώνες μέσω της ηλεκτροφόρησης πηκτωμάτων.

Εικόνα 30 [40]
Ειδικής μεθυλίωσης PCR (MSP)(Εικόνα 31) χρησιμοποιείται για να ανιχνεύσει τη μεθυλίωση CpG νήσων στο γενωμικό DNA. Το DNA αντιμετωπίζεται αρχικά με δισουκφιδικό νατρίο, που μετατρέπει μη μεθυλιωμένες βάσεις κυτοσίνης σε ουρακίλη, που αναγνωρίζεται από τους PCR εκκινητές σαν θυμίνη. Δύο pcr αντιδράσεις πραγματοποιούνται έπειτα στο τροποποιημένο DNA, με χρήση sets ίδιων εκκινητών εκτός από κάθε CpG νήσους μέσα στις αλληλουχίες του εκκινητή. Σε αυτά τα σημεία, ένα σύνολο εκκινητών αναγνωρίζει DNA με κυτοσίνες για να ενισχύσει το μεθυλιωμένο DNA, και ένα σύνολο αναγνωρίζει DNA με την ουρακίλη ή θυμίνη για να ενισχύσει μη μεθυκιωμένο DNA. MSP με χρήση qPCR μπορεί επίσης να εκτελεσθεί για να λάβει ποσοτικές παρά ποιοτικές πληροφορίες για τη μεθυλίωση.

Εικόνα 31 [41]
Στρατηγική για την methylation-specific PCR με βάση το σχέδιο μεθυλίωσης της FMR1 CpG νήσου. Οι μεθυλιωμένες κυτοσίνες παρουσιάζονται με κόκκινο και χαρακτηρίζονται ως αστερίσκοι. Οι μη μεθυλιωμένες κυτισίνες μετατρέπονται σε κατάλοιπα ουρακίλης (που παρουσιάζονται με μπλε) με δισουλφιδική επεξεργασία. Αυτό το διάγραμμα είναι βασισμένο στη δημοσίευση από τον Zeschnigk και άλλους. (1997) για τη διάγνωση των Angelman και Prader-Willi συνδρόμων με MSP.[41]

Εικόνα 32 [21]
Διάγραμμα ροής που περιγράφει την εφαρμογή της in situ RT-PCR (Εικόνα 32) σε μικροσυστοιχίες ιστού. Βήμα 1: Προετοιμασία φωτογραφικών διαφανειών. Τμήματα που αποπαραφινοποιήθηκαν με τα πλυσίματα ξυλολίων, και κυτταρικά υλικά εμποτίστηκαν από την ειδική πρωτεϊνάση πέψης K; Βήμα 2: ενίσχυση αντίστροφα-μεταγραφόμενου cDNA με εσωνίων-μέτρηση PCR εκκινητές (οριζόντια βέλη) προστιθέμενος σε ένα απλό μίγμα αντίστροφης, τρανσκρυπτάσης. Η πολυμεράση τα δεοξυριβονουκλεοτίδια ενωμένα με το digoxygenin -σημαδεμένο dUTP (μαύρα ovals); Βήμα 3: απεικόνιση των PCR προϊόντων με σύνδεση digoxygenin-ειδικών με χρυσό-σημαδεμένων αντισωμάτων (κίτρινα σημεία), ακολουθούμενη με ασημί πυρήνωση γύρω από τους δεσμούς των χρυσών μορίων (γκρίζες ημισέληνοι). [21]
AFLP PCR
Μια πρωτότυπη τεχνική DNA δακτυλοσκοπίας αποκαλούμενη AFLP περιγράφεται. Η AFLP τεχνική είναι βασισμένη στην εκλεκτική PCR ενίσχυση από τον περιορισμό τμημάτων από ένα σύνολο γενωμικού DNA. Η τεχνική περιλαμβάνει τρία βήματα: (i) περιορισμός του DNA και σύνδεση με ολιγονουκλεοτίδια προσαρμογείς, (ii) εκλεκτική ενίσχυση του συνόλου των τμημάτων περιορισμού, και (iii) ανάλυση σε πηκτή των ενισχυμένων τμημάτων. PCR ενίσχυση των τμημάτων περιορισμού επιτυγχάνεται με χρήση προσαρμογέα και περιοχών ακολουθίας περιορισμού σαν περιοχές στόχων για υβριδισμό του εκκινητή. Η εκλεκτική ενίσχυση επιτυγχάνεται με χρήση εκκινητών που επεκτείνουν τα τμήματα περιορισμού, με ενίσχυση μόνο εκείνων των τμημάτων στα οποία οι επεκτάσεις των εκκινητών τεριάζουν στα νουκλεοτίδια που πλαισιώνουν τις περιοχές περιορισμού. Χρησιμοποίηση αυτής της μεθόδου, σύνολα τμημάτων περιορισμού μπορούν να απεικονιστούν με PCR χωρίς γνώση της ακολουθίας των νουκλεοτιδίων. Η μέθοδος επιτρέπει την ειδική ομο-ενίσχυση σε μεγάλους αριθμούς τμημάτων περιορισμού. Ο αριθμός των τμημάτων που μπορεί να αναλυθεί ταυτόχρονα, εντούτοις, είναι εξαρτώμενος από την ευαισθησία του συστήματος ανίχνευσης. Χαρακτηριστικά 50-100 περιορισμού τμήματα ενισχύονται και ανιχνεύονται στην αποδιάταξη σε πηκτώματα πολυακρυλαμιδίου. Η AFLP η τεχνική παρέχει μια πρωτότυπη και πολύ ισχυρή τεχνική δακτυλοσκόπησης του DNA για DNAs με οποιαδήποτε προέλευση ή πολυπλοκότητα.[42]


Εικόνα 33 Σύνδεση των προσαρμοστών σε AFLP-PCR πειράματα

Εικόνα 34 AFLP-PCR με προσαρμογείς τεριάσματος εκκινητών και με ένζυμα περιορισμού αναγνώρισης ακολουθιών.

Εικόνα 35 AFLP-PCR, με προσαρμογείς τεριάσματος εκκινητών, με ένζυμα περιορισμού αναγνώρισης ακολουθιών και με περιεκτικότητα σε εκλεκτικές τελικές βάσεις στο 3' άκρο. Η χρήση εκλεκτικών εκκινητών θα μειώσει τον αριθμό από τις ζώνες που ενισχύονται [43]
Πρόσφατες εξελίξεις στις PCR τεχνικές
· Μια πιό πρόσφατη μέθοδος που αποκλείει έναν κύκλο θερμοκρασίας, αλλά χρησιμοποιεί ένζυμα, είναι η ελικάση - εξαρτώμενη ενίσχυση (HDA).
· TAIL-PCR, αναπτύχθηκε από τον Liu και άλλους. το 1995, είναι θερμική ασυμμετρική συμπλεγμένη PCR.
· Meta-PCR, αναπτύχθηκε από τον Andrew Wallace, επιτρέπει την βελτιστοποίηση της ενίσχυσης και την άμεση ανάλυση μιας ακολουθίας από σύνθετα γονίδια.
Helicase-dependent amplification (HDA)
Η Ελικάση-εξαρτώμενη ενίσχυση (HDA) είναι μια μέθοδος για in vitro DNA ενίσχυση όπως η PCR.(Εικόνα 36)
Το μειονέκτημα της PCR είναι ότι καταναλώνει πολύ χρόνο με το ξετύλιγμα του δίκλωνου DNA με τη θέρμανση των απλών κλώνων (μια διαδικασία αποκαλούμενη αποδιάταξη) και αντιγραφή των απλών κλώνων για να δημιουργήσουν νέο δίκλωνο DNA (σύνθεση). Αντί αυτών των thermocycles, η HDA μιμείται φυσικές μεθόδους αντιγραφής του DNA με τη χρησιμοποίηση ελικάσης (ένα ένζυμο) που αποδιατάσσει το DNA σε μια σταθερή θερμοκρασία

Εικόνα 36 Σχηματικό διάγραμμα της HDA. Δύο συμπληρωματικοί DNA κλώνοι παρουσιάζεται ως δύο γραμμές: ο παχύς είναι ο πάνω κλώνος και ο λεπτός είναι ο κάτω κλώνος 1: Μια ελικάση (μαύρο τρίγωνο) χωρίζει τους δύο συμπληρωματικούς DNA κλώνους, που είναι συνδεδεμένοι με SSB (γκρίζοι κύκλοι). 2: Εκκινητές (γραμμές με τους δείκτες των βελών) υβριδιοποιούν στην περιοχή στόχων ssDNA πρότυπο. 3: Μια DNA πολυμεράση (τετράγωνα με τα σχέδια μωσαϊκών) επεκτείνει τους εκκινητές υβριδοποιόντας το πρότυπο DNA. 4: Ενισχυμένα προϊόντα εισάγονται στον επόμενο κύκλο της ενίσχυσης.[ω44]
Τα πλεονεκτήματα της HDA έναντι της PCR είναι η αυξανόμενη ταχύτητα, μειωμένες δαπάνες και η προοπτική δημιουργίας μιας χειροπιαστής DNA διαγνωστικής συσκευής. Το μειονέκτημα είναι ότι η HDA ακόμα απαιτεί ένα σχετικά μεγάλο ποσό DNA, έτσι αυτή πιθανώς να μην χρησιμοποιηθεί σε περιπτώσεις όπου απαιτείται υψηλή ευαισθησία. [45]
|
|


Εικόνα 37 [46]
Η TAIL είναι μια σειρά αντιδράσεων που προορίζονται να χαρτογραφήσουν που ένα T-DNA (μεταφορικό DNA) έχει παρεμβληθεί μέσα στο γονιδίωμα. Τα κύρια συστατικά από τις 3 αντιδράσεις είναι AD (Αυθαίρετοι εκφυλισμένοι) εκκινητές, ορόσημα εκκινητές, και DNA από T-DNA γραμμές που πρόκειται να χαρτογραφηθούν. AD εκκινητές είναι εκφυλισμένοι εκκινητές που υβριδοποιούν σε όλο το γονιδίωμα. Τα ορόσημα εκκινητές είναι ειδικοί για το αριστερό και δεξιό όριο του T-DNA. Από την αρχική αντίδραση στην τριτογενή, οι εκκινητές ορόσημα τοποθετούνται πιό κοντά στην άκρη του T-DNA. Γί’αυτό μια "μετατόπιση" είναι ορατή κατά την τρέξιμο σε ένα πηκτώματος κατά τις δευτερογενείς και τριτογενείς αντιδράσεις έπειτα από την κάθε μία. Το ποσοστό επιτυχίας της TAIL-PCR ποικίλλει, εξαρτάται δε από το ποσό των DNA,δειγμάτων ,των AD εκκινητών, και των ορόσημων εκκινητών που χρησιμοποιούνται.[47]

Εικόνα 38 Σχηματική αναπαράσταση της TAIL PCR. Αυτή η στρατηγική περιλαμβάνει τρείς διαδοχικούς κύκλους PCR, διενεργηθέντες με ένα σύνολο τριών nested T-DNA ειδικών εκκινητών (SP1, SP2 and SP3) και έναν μικρό, αυθαίρετο εκκινητής. Οι θέσεις των τριών nested εκκινητών στην T-DNA περιοχή υποδεικνύονται από τα μπλε, κόκκινα και πράσινα βέλη αντίστοιχα. AD είναι ο αυθαίρετος εκφυλισμένος εκκινητής υποδεδειγμένος από το πορφυρό βέλος. Η T-DNA περιοχή πλαισίωσης υποδεικνύεται από την πράσινη γραμμή. [48] |
Meta-PCR
Υπόβαθρο και αρχές
Η Meta-PCR είναι μια μέθοδος για τη δημιουργία ενός συνθετικού μορίου DNA συμπεριλαμβάνοντας οποιονδήποτε συνδυασμό PCR επαύξησης του DNA για κάθε περαγγελία. Πραγματοποιείται σε έναν απλό σωλήνα και αποτελείται από δύο χωριστά βήματα PCR χωρισμένα με έναν κύκλο τήξης-πήξης για να αφαιρέσει την υπόλοιπη δραστηριότητα των πολυμεράσεων. Meta-PCR μπορεί να χρησιμοποιηθεί για να αυξηθεί η ρυθμοαπόδοση των μεθόδων ανίχνευσης μεταλλαγής. Το μέσο μέγεθος ενός εξονίου σε ένα γονίδιο ανθρώπου είναι μόνο 150bp. Οι περισσότερες τεχνικές ανίχνευσης μεταλλαγής είναι σε θέση της ανάλυσης πολύ μεγαλύτερων τμημάτων DNA. Συνδυασμός διάφορων μεμονωμένων εξονίων από κοινού σε ένα από DNA τμήμα μπορούν έτσι να αυξήσουν την ρυθμοαπόδοση με τη μείωση της καθοδικής επεξεργασίας και των αναλυτικών βημάτων. Έχουμε χρησιμοποιήσει Meta-PCR συνδεδεμένη με άμεση αλληλούχηση για να βελτιώσει τη ρυθμοαπόδοση της ανίχνευσης μεταλλαγής. Η αρχική Meta-PCR ενίσχυση είναι μια multiplex PCR με χρήση ζευγαριών εκκινητών στον περιορισμό των συγκεντρώσεων (0.04µM κάθε ένας) που έχουν συμπληρωματικά 5' άκρα συνδεδεμένα με βάσεις αδενίνης σε συγκεκριμένες

Εικόνα 39: Πρωταρχική 1 Meta-PCR: multiplex ενίσχυση από μεμονωμένα εξόνια από γενωμικό πρότυπο DNA Κατά τη διάρκεια των αρχικών καταστάσεων τα αρχικά Meta-PCR μεμονωμένα τμήματα απλώς ενισχύονται. Κατά τη διάρκεια αυτής της φάσης ο συμπληρωματικός συνδετήρας ενσωματώνει τα τμήματα. Προς το τέλος της αρχικής Meta-PCR ενίσχυσης οι εκκινητές εξαντλούνται και επέκταση επικάλυψης μπορεί να εμφανιστεί, παράγοντας κατά συνέπεια μικρές ποσότητες από πλήρους μήκους συνθετικά προϊόντα (Εικόνα 40).
Η διαδικασία σύνδεσης είναι αυτό-συγκεντρωμένη και οδηγείται από το δεύτερο νόμο της θερμοδυναμικής με επακόλουθο κέρδος της εντροπίας του συστήματος συνολικά.

Εικόνα 40: Μετά 1 Meta-PCR: τα εξόνια ευθυγραμμίζονται και συγκεντρώνονται μέσω των 5 "συμπληρωματικών συνδετικών ουρών. Με την ολοκλήρωση της αρχικής Meta-PCR αντίδρασης παγώνει στους -20οC για τουλάχιστον 1 ώρα προκειμένου να αδρανοποιήσει την υπόλοιπη δραστηριότητα των πολυμερασών. Αυτό εξασφαλίζει ότι η ‘Hot Start’ που συστήνεται και για τις δύο ενισχύσεις, την αρχική και την δευτεροβάθμια Met-PCR είναι πλήρης και εξασφαλίζει έτσι υψηλή εξειδίκευση αντίδρασης. Η δευτεροβάθμια Meta-PCR ενίσχυση περιλαμβάνει έναν δεύτερο κύκλο PCR ενίσχυση με χρήση 1/10 όγκο από το πάγωμα / ξεπάγωμα της αρχικής Meta-PCR αντίδρασης ως πρότυπο. Ένα απλό ζευγάρι εσωτερικά των εκκινητών της nested PCR επαν-ενισχύει μια μικρή ποσότητα πλήρους μήκους συνδεμένου προϊόντος όποιος έχει προκύψει στο τέλος από την αρχική αντίδραση (Εικόνα 41).
Έχουμε σχεδιάσει εσωτερικά nested εκκινητές για να κατέχουν μία καθολική

Εικόνα 41: 2 Meta-PCR: Πλήρως συγκεντρωμένα Meta-PCR τμήματα ενισχύονται σε υψηλό αριθμό αντιγράφων με χρήση εκκινητή nested εσωτερικών ζευγών. [45]
PCR μπορεί να χρησιμοποιηθεί για ένα ευρύ φάσμα πειραμάτων και αναλύσεων. Μερικά παραδείγματα αναφέρονται κατωτέρω.[49]
Γενετική δακτυλοσκοπία είναι μια δικανική τεχνική που χρησιμοποιείτα για να προσδιοριστεί ένα πρόσωπο με τη σύγκριση του DNA του ή της από ένα δεδομένο δείγμα. Ένα παράδειγμα είναι :το αίμα από τον τόπο του εγκλήματος μπορεί να συγκριθεί γενετικά με το αίμα από έναν ύποπτο. Το δείγμα μπορεί να περιέχει μόνο ένα μικροσκοπικό ποσό DNA (αποκτηθέν από μια πηγή όπως το αίμα, σπέρμα, σάλιο, τρίχα , ή άλλο οργανικό υλικό). Θεωρητικά, χρειάζεται ένας απλός κλώνος. Κατ' αρχάς, σπάζουμε το DNA δείγμα σε τμήματα, κατόπιν τα ενισχύουμε χρησιμοποιώντας PCR. Τα ενισχυμένα τμήματα μετά χωρίζουν χρησιμοποίηση της ηλεκτροφόρησης πηκτωμάτων. Το γενικό σχεδιάγραμμα DNA τμημάτων καλείται δακτυλικό αποτύπωμα DNA. Δεδομένου ότι υπάρχει μια πολύ μικρή δυνατότητα εκείνα τα δύο άτομα μπορούν να έχουν τις ίδιες ακολουθίες (μία σε αρκετά εκατομμύρια), η τεχνική είναι αποτελεσματικότερη στην αθώωση ενός ύποπτου από ότι η παρουσίαση αποδείξεων για την ενοχή ενός ύποπτου.[49]

Εικόνα 42 Ηλεκτροφόρηση από PCR- ενισχυμένα DNA τμήματα. (1) Πατέρας. (2) Παιδί. (3) Μητέρα. Το παιδί έχει κληρονομήσει μερικά, αλλά όχι όλο το γενετικό αποτύπωμα από κάθε έναν από τους γονείς του, που δίνει σ’αυτόν ένα νέο, μοναδικό γενετικό αποτύπωμα. [2]
Αν και αυτά τα προκύπτοντα "γενετικά αποτυπώματα" είναι μοναδικά (εκτός από τους μονογενείς δίδυμους), γενετικές σχέσεις, παραδείγματος χάριν, γονέα - παιδιού ή αμφιθαλών, μπορούν να καθοριστούν από δύο ή περισσότερα γενετικά αποτυπώματα, που μπορούν να χρησιμοποιηθούν για τις δοκιμές πατρότητας. Μια παραλλαγή αυτής της τεχνικής μπορεί επίσης να χρησιμοποιηθεί για να καθορίσει τις εξελικτικές σχέσεις μεταξύ των οργανισμών.[49]
Η ανίχνευση κληρονομικών ασθενειών σε ένα δεδομένο γονιδίωμα είναι μια μακροχρόνια και δύσκολη διαδικασία, που μπορεί να συντομευθεί σημαντικά με τη χρησιμοποίηση PCR. Κάθε εν λόγω γονίδιο μπορεί εύκολα να ενισχυθεί μέσω PCR με τη χρησιμοποίηση τον κατάλληλου εκκινητή και έπειτα με αλληλούχηση να ανιχνευθούν μεταλλαγές.Προερχόμενες από ιό ασθένειες, επίσης, μπορούν να ανιχνευτούν με χρήση PCR μέσω της ενίσχυσης του προερχόμενου από τον ιό DNA. Αυτή η ανάλυση είναι δυνατόν να γίνει σωστά μετά από τη μόλυνση, που μπορεί να είναι από αρκετές ημέρες έως αρκετούς μήνες πριν εμφανιστείτούν τα πραγματικά συμπτώματα. Τέτοιες έγκαιρες διαγνώσεις παράσχουν στους παθολόγους σημαντική βοήθεια στην επεξεργασία.[49]
Η κλωνοποίηση ενός γονιδίου, (Εικόνα 43) για να μην συγχέεται με την κλωνοποίηση ολόκληρου του οργανισμού, περιγράφει τη διαδικασία, από την απομόνωση ενός γονιδίου από έναν οργανισμό και την εισαγωγή έπειτα αυτού σε έναν άλλο οργανισμό (που τώρα καλείται γενετικά τροποποιημένος οργανισμός (GMO)). PCR χρησιμοποιείται συχνά για να ενισχύσει το γονίδιο, που μπορεί έπειτα να εισαχθεί σε ένα φορέα κλωνοποίησης (φορέας κλωνοποίησης είναι ένα τμήμα DNA που "φέρνει" το γονίδιο GMO) όπως το πλασμίδιο (ένα κυκλικό μόριο DNA). Το DNA μπορεί έπειτα να μεταφερθεί σε έναν οργανισμό (GMO) όπου το γονίδιο και το προϊόν του μπορούν να μελετηθούν περισσότερο. Έκφραση ενός κλωνοποιημένου γονιδίου (όταν σε ένα γονίδιο εκφράζεται το προϊόν γονιδίων (συνήθως πρωτεϊνη ή RNA) παράγεται από GMO) μπορεί επίσης να είναι ένας τρόπος που παράγει μαζικά χρήσιμες πρωτεϊνες, παραδείγματος χάριν φάρμακα ή ένζυμα σε βιολογικές σκόνες πλύσης. Η ενσωμάτωση μιας ετικέττας συγγένειας σε μιά ανασυνδυαζόμενη πρωτεϊνη θα παραγάγει μιά μεικτή πρωτεϊνη που μπορεί να προσδιοριστεί ευκολότερα με χρωματογραφία συγγένειας .

Εικόνα 43 Κλωνοποίηση ενός γονιδίου που χρησιμοποιεί ένα πλασμίδιο.[50]
(1) Χρωμοσωμικό DNA από τον οργανισμό A. (2) PCR. (3) Πολλαπλάσια αντίγραφα από ένα απλό γονίδιο από τον οργανισμό A. (4) Εισαγωγή του γονιδίου σε ένα πλασμίδιο. (5) Πλασμίδιο με το γονίδιο από τον οργανισμό A. (6) Εισαγωγή του πλασμιδίου στον οργανισμό B. (7) Πολλαπλασιασμός ή έκφραση του γονιδίου, αρχικά από τον οργανισμό A, και εμφάνιση στον οργανισμό B.
Μεταλλαξιγένεση είναι ένας τρόπος να προκαλείς αλλαγές στην ακολουθία των νουκλεοτιδών του DNA. Υπάρχουν καταστάσεις στις οποίες ένας ενδιαφέρεται για μεταλλαγμένα (αλλαγμένα) αντίγραφα από ένα δεδομένο DNA κλώνο, παραδείγματος χάριν, κατά την προσπάθεια να αξιολογηθεί η λειτουργία από ένα γονίδιο ή σε in-vitro εξέλιξη πρωτεϊνης (επίσης γνωστή ως κατευθυνόμενη εξέλιξη). Οι μεταλλαγές μπορούν να εισαχθούν σε αντιγραμμένες DNA ακολουθίες με δύο πλήρως διαφορετικές PCR διαδικασίες. Περιοχή-κατευθυνμένη μεταλλαξιγένεση επιτρέπει στον πειραματιστή να εισαγάγει μια μεταλλαγή σε μια συγκεκριμένη θέση στο DNA κλώνο. Συνήθως, η επιθυμητή μεταλλαγή ενσωματώνεται στους εκκινητές με χρήση ενός PCR προγράμματος. Τυχαία μεταλλαξιγένεση, αφ' ετέρου, είναι βασισμένη στη χρήση μιας πολυμεράσης επιρρεπούς σε λάθη στην pcr διαδικασία. Στην περίπτωση της τυχαίας μεταλλαξιγένεσης, η θέση και η φύση από τις μεταλλαγές δεν μπορεί να ελεγχθεί. Μια εφαρμογή της τυχαίας μεταλλαξιγένεσης είναι να αναλύσει τις σχέσεις δομή-λειτουργίας από μια πρωτεϊνη. Με τυχαία μετάλλαξη μπορεί κάποιος να αλλάξει μια DNA ακολουθία, μπορεί να συγκρίνει την προκύπτουσα πρωτεϊνη με την αρχική και να καθορίσει τη λειτουργία κάθε μέρους της πρωτεϊνης.[2]
Με χρήση PCR, είναι δυνατό να αναλυθεί το DNA που είναι χιλιάδες έτη παλαιό. Οι PCR τεχνικές έχουν χρησιμοποιηθεί επιτυχώς στα ζώα, όπως σε ένα σαράντα-χιλιάδων-ετών- μαμμούθ, και επίσης στο ανθρώπινο DNA, στην έκταση εφαρμογών από την ανάλυση σε Αιγυπτιακές μούμιες μέχρι τον προσδιορισμό των Ρώσων Τσάρων.[2]
Μέσω της χρήσης της allele-specific PCR, κάποιος μπορεί εύκολα να καθορίσει ποιο αλληλόμορφο γονίδιο μιας μεταλλαγής ή πολυμορφισμό έχει ένα άτομο. Εδώ, ένας από τους δύο εκκινητές είναι κοινός, και θα υβρίδιζε σε μικρή απόσταση μακρυά από τη μεταλλαγή, ενώ άλλος θα υβρίδιζε δεξιά από την παραλλαγή. Το 3' άκρο του αλληλομόρφου-ειδικού εκκινητή τροποποιείται, για να υβριδίσει μόνο εάν ταιριάζει με ένα από τα αλληλόμορφα γονίδια. Εάν η μεταλλαγή ενδιαφέροντος είναι μια T ή C απλού νουκλεοτιδίου πολυμορφισμός (T/C SNP), κάποιος θα χρησιμοποιούσε δύο αντιδράσεις, μία που να περιέχει έναν εκκινητή που τελειώνει με T, και μία άλλη με τελείωμα σε C. Ο κοινός εκκινητής θα ήταν ο ίδιος. Μετά από την PCR, αυτά τα δύο σύνολα αντιδράσεων θα έτρεχαν σε ένα πήκτωμα αγαρόζης, και το σχέδιο ζωνών θα μας έλεγαν εάν το άτομο είναι ομοζυγώτης T, ομοζυγώτης C, ή ετερογυζυγώτικός T/C. Αυτή η μεθοδολογία έχει διάφορες εφαρμογές, όπως η ενίσχυση ορισμένων απλότυπων (όταν ορισμένα αλληλόμορφα γονίδια σε 2 ή περισσότερα SNPs εμφανίζονται μαζί στο ίδιο χρωμόσωμα.[2]
Ερευνητές έχουν χρησιμοποιήσει παραδοσιακή PCR σαν τρόπο να υπολογίσουν αλλαγές στο ποσό από της έκφραση των γονιδίων. Το RNA είναι το μόριο στο οποίο το DNA μεταγράφεται πριν από την κατασκευή μιας πρωτεϊ'νης, και εκείνοι οι κλώνοι του RNA που φέρουν τις οδηγίες για την πρωτεϊνική ακολουθία είναι γνωστοί ως αγγελιοφόρο RNA (mRNA). Μόλις απομονωθεί το RNA μπορεί να γίνει αντίστροφη μεταγραφή πάλι σε DNA (συμπληρωματικό DNA γνωστό σαν cDNA), και σε κάποιο σημείο της παραδοσιακής PCR μπορεί να εφαρμοστεί για να ενισχύσει το γονίδιο. Αυτή η μεθοδολογία καλείται RT-PCR. Στις περισσότερες περιπτώσεις εάν υπάρχει περισσότερο αρχικό υλικό (mRNA) από ένα γονίδιο κατά τη διάρκεια της PCR περισσότερα αντίγραφα του γονιδίου θα παραχθούν. Όταν τα προϊόντα της PCR οργανώνονται σε ένα πήκτωμα αγαρόζης μια ζώνη, που αντιστοιχεί σε ένα γονίδιο, θα εμφανιστεί μεγαλύτερη στο πήκτωμα (σημειώστε ότι αν η ζώνη παραμένει στην ίδια θέση σχετικά με τη κλίμακα, θα εμφανιστεί ακριβώς παχύτερη ή φωτεινότερη). Με το τρέξιμο των δειγμάτων του ενισχυμένου cDNA από διαφορετικά αντιμετωπισμένους οργανισμούς κάποιος μπορεί να πάρει μια γενική ιδέα για το ποιό δείγμα του γονιδίου που ενδιαφέρεται εκφράζεται περισσότερο. Μια quantative RT-PCR μέθοδος έχει αναπτυχθεί, και καλείται Real-time PCR .[2]
Η PCR τεχνική ήταν κατοχυρωμένη με δίπλωμα ευρεσιτεχνίας από την Cetus Corporation, όπου ο Mullis εργάστηκε όταν εφηύρε η τεχνική το 1983. Η πολυμεράση Taq το ένζυμο καλύπτεται επίσης από τα διπλώματα ευρεσιτεχνίας. Έχουν υπάρξει αρκετές υψηλού προφίλ αγωγές σχετικές με την τεχνική, συμπεριλαμβανομένου μιας ανεπιτυχούς δίκης παρουσιασμένης από τον DuPont. Η φαρμακοβιομηχανία Hoffmann-La Roche αγόρασε τα δικαιώματα στα διπλώματα ευρεσιτεχνίας το 1992 και κρατά αυτήν την περίοδο εκείνα που προστατεύονται ακόμα.
Μια σχετική μάχη διπλωμάτων ευρεσιτεχνίας πέρα από το ένζυμο Taq πολυμεράση βρίσκεται ακόμα σε εξέλιξη σε αρκετά θέματα σε όλο τον κόσμο μεταξύ της Roche και της Promega. Κατά τρόπο ενδιαφέροντα, φαίνεται που πιθανώς τα νομικά επιχειρήματα θα επεκταθούν πέρα από τη ζωή για τα διπλώματα ευρεσιτεχνίας για την αρχική pcr και Taq πολυμεράση, που έληξε τις 28 Μαρτίου, 2005.[51]
2η παρουσίαση σε PowerPoint
[1] The history of PCR: Smithsonian Institution Archives, Institutional History Division. Retrieved 24 June 2006.
[2] http://en.wikipedia.org/wiki/Polymerase_chain_reaction
[3] Mullis, Kary (1998). Dancing Naked in the Mind Field. New York: Pantheon Books. ISBN 0-679-44255-3.
[4] http://content.answers.com/main/content/wp/en/thumb/a/aa/200px-Pcr_machine.jpg
[5] Sambrook, Joseph; and David W. Russell (2001). Molecular Cloning: A Laboratory Manual, 3rd ed., Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press. ISBN 0-87969-576-5.
[6] Μπουρτζής Κ., Εργαστηριακές Ασκήσεις Βιοχημείας και Μοριακής Βιολογίας, Πανεπιστήμιο Ιωαννίνων ,2002, σελ. 31-32.
[7]http://www.cs.colorado.edu/ugrad/seniorproject/projects/shots/geneact/primer.png
[8]http://www.amath.washington.edu/~qian/TheScientist/TheScientists_files/geyser.jpg
[9] http://www.stratagene.com/products/showCategory.aspx?catId=116
[10] http://content.answers.com/main/content/wp/en/5/56/Pcr.png
[11]http://www.oceanexplorer.noaa.gov/explorations/04etta/background/dna/media/dna_2.html
[12] Πέτρου Παναγιώτα, ΣΥΣΤΟΙΧΙΕΣ ΜΙΚΡΟΚΗΛΙ∆ΩΝ ΟΛΙΓΟΝΟΥΚΛΕΟΤΙ∆ΙΩΝ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΜΕΤΑΛΛΑΞΕΩΝ, Εργαστήριο Ανοσοαναλύσεων / Ανοσοαισθητήρων, Ινστιτούτο Ραδιοϊσοτόπων ––Ραδιοδιαγνωστικών Προϊόντων, ΕΚΕΦΕ «∆ηµόκριτος», Θερινό σχολείο, 11-22 Ιουλίου, 2005.
[13] Pavlov AR, Belova GI, Kozyavkin SA, Slesarev AI (2002). "Helix-hairpin-helix motifs confer salt resistance and processivity on chimeric DNA polymerases". Proc Natl Acad Sci. 99: 3510-13515. PMID 12368475.
[14] Demidov VV (2002). "A happy marriage: advancing DNA polymerases with DNA topoisomerase supplements". Trends Biotechnol. 20: 491. DOI:10.1016/S0167-7799(02)02101-7.
[15] Pavlov AR, Pavlova NV, Kozyavkin SA, Slesarev AI (2004). "Recent developments in the optimization of thermostable DNA polymerases for efficient applications". Trends Biotechnol. 22: 253-260. PMID 15109812.
[16] Pavlov AR, Pavlova NV, Kozyavkin SA, Slesarev AI (2004). “Thermostable Chimeric DNA Polymerases with High Resistance to Inhibitors”, DNA Amplification: Current Technologies and Applications. Horizon Bioscience, pp. 3-20. ISBN 0-9545232-9-6.
[17] Forterre P (2006). "DNA topoisomerase V: a new fold of mysterious origin". Trends Biotechnol. 24: 245-247. PMID 16650908.
[18] Pavlov AR, Pavlova NV, Kozyavkin SA, Slesarev AI (2006). “Thermostable DNA Polymerases for a Wide Spectrum of Applications: Comparison of a Robust Hybrid TopoTaq to other enzymes”, Kieleczawa J: DNA Sequencing II: Optimizing Preparation and Cleanup. Jones and Bartlett, pp. 241-257. ISBN 0-7637338-3-0.
[19] Taneja B, Patel A, Slesarev A, Mondragon A (2006). "Structure of the N-terminal fragment of topoisomerase V reveals a new family of topoisomerases". EMBO J. 25: 398-408. PMID 16395333.
[20] http://www.ars.usda.gov/pandp/docs.htm?docid=11318
[21] http://bmc.ub.uni-potsdam.de/1477-3155-1-3/F2.htm
[22] http://210.212.212.7:9999/PHP/SILKSAT/img/issr_diagram.png
[23] http://www.eng.buffalo.edu/~coffroth/ISSR-PCR.html
[24] Nagaraju J.; Kathirvel M.; Subbaiah E.V.; Muthulakshmi M.; Kumar L.D., FISSR-PCR: a simple and sensitive assay for highthroughput genotyping and genetic mapping, Molecular and Cellular Probes, Volume 16, Number 1, February 2002, pp. 67-72(06), Academic Press.
[25] http://www.envfor.nic.in/report/0203/chap-07.htm
[26] Shu-Mei Dai, Hsiu-Hua Chen, Cheng Chang, Arthur D. Riggs & Steven D. Flanagan, Ligation-mediated PCR for quantitative in vivo footprinting, Nature Biotechnology 18, 1108 - 1111 (2000) doi:10.1038/80323
[27] http://www.bio.uu.nl/mg/pd/research/index.html
[28]http://www.bx.psu.edu/~ross/workmg/Isolating_analyzing_genes_files/image109.jpg
[29] Kenji Abe, Direct PCR from Serum, Application to Viral Genome Detection, Methods in Molecular Biology, Vol. 226: PCR Protocols, Second EditionEdited by: J. M. S. Bartlett and D. Stirling, Humana Press Inc., Totowa, NJ, p.164.
[30] http://startrek.ccs.yorku.ca/~pjohnson/AssemblyPCRProtocol.html
[31] http://www.promega.co.jp/jp/jp_tech/FAQs/Q&A_Plexor.html
[32] http://www.iba-go.com/images/naps/rt_pcr.gif
[33] Francisco Vega, Jeffrey L Medeiros, Rayjalakshmi Luthra: Taqman PCR In The Detection And Quantification Of Chromosomal Translocations In Follicular Lymphoma, Mantle Cell Lymphoma And Chronic Myeloid Leukemia: Taqman Real-time PCR Assay. The Internet Journal of Pathology. 2002. Volume 2 Number 1.
[34] http://www.takaramirusbio.com/html/PCR_RTPCR_INTRO.html
[35] http://www.superarray.com/newsletter/validating.html
[36] http://www.mcmaster.ca/inabis98/higuchi/sung0691/four.html
[37] http://www.takaramirusbio.com/images/PCR_IMD_PROFILE.gif
[38] http://www.epibio.com/item.asp?ID=401
[39] http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=hmg.figgrp.2574
[40] http://www.wiley.com/legacy/products/subject/life/genetics/fig610.jpg
[41] http://www.scielo.br/img/fbpe/gmb/v22n2/html/0276f1.htm
[42] P Vos, R Hogers, M Bleeker, M Reijans, T van de Lee, M Hornes, A Frijters, J Pot, J Peleman, and M Kuiper, Keygene N.V., AFLP: a new technique for DNA fingerprinting.Nucleic Acids Res. 1995 November 11; 23(21): 4407–4414.
[43] Zabeau, M and P. Vos. 1993. Selective restriction fragment amplification: a general method for DNA fingerprinting. European Pattent Office, publication 0 534 858 A1, bulletin 93/13.
[44] Myriam Vincent, Yan Xu & Huimin Kong, Helicase-dependent isothermal DNA amplification, EMBO reports 5, 8, 795–800 (2004) doi:10.1038/sj.embor.7400200, Published online: 9 July 2004
[45] NATIONAL GENETICS REFERENCE LABORATORY, Mutation Scanning incomplex genes usingMeta-PCR and DNAsequencing, (Manchester), November 2004, p.3-5.
[46] http://www.pyc.pku.edu.cn/database/pcr.html
[47] http://preuss.bsd.uchicago.edu/protocols/tail.html
[48] http://www.ejbiotechnology.info/content/vol8/issue1/full/4/f5.html
[49] http://en.wikipedia.org/wiki/Polymerase_chain_reaction
[50] http://upload.wikimedia.org/wikipedia/commons/0/07/Pcr_clone.png
[51] Advice on How to Survive the Taq Wars ¶2: GEN Genetic Engineering News Biobusiness Channel: Article. May 1 2006 (Vol. 26, No. 9).
13. Μέθοδοι ανάλυσης μεταλλάξεων χωρίς προσδιορισμό των νουκλεοτιδικών αλληλουχιών: αρχή μεθόδων, πειραματικά πρωτοκόλλα, διδακτικά σχήματα.
Η μεταλλαγή στα γονίδια είναι υπεύθυνη για διάφορες γενετικές διαταραχές. Ο προσδιορισμός αυτών των μεταλλαγών απαιτεί νέες μεθόδους ανίχνευσης. Εντατική έρευνα κατά τη διάρκεια των ετών έχει οδηγήσει σε διάφορες νέες τεχνικές ανίχνευσης, μερικές από τις όποιες είναι εξειδικευμένες, όπως για παράδειγμα για τον καρκίνο του μαστού. Αυτές οι αναλύσεις γίνονται λαμβάνοντας υπόψη, το ιστορικό του καρκίνου, του ασθενή και το οικογενειακό ιστορικό
· Allele Specific Oligonucleotides (ASO).
Υβριδισμός με ειδικά ολιγονουκλεοτίδια.
· Protein Truncation Test (PTT).
Πρωτεϊνική δοκιμή αποκοπής.
· Single
Πολυμορφισμός διαμόρφωσης μονόκλωνης αλυσίδας.
· Nucleotide sequencing.
Αλληλούχηση νουκλεοτιδίων.
· Denaturing Gradient Gel Electrophoresis (DGGE).
Ηλεκτροφόρηση σε πηκτή με κλίση αποδιατακτικού.
· Heteroduplex analysis.
Ετερόδιπλου μορίου ανάλυση.
· DNA microarray technology.
Τεχνολογία DNA μικρομωσαϊκών.
· Temperature gradient gel electrophoresis (TGGE).
Ηλεκτροφόρηση σε πηκτή βαθμιδωτής θερμοκρασίας (παραλλαγή της DGGE).
· DENATURING HIGH-PERFORMANCE LIQUID CHROMATOGRAPHY (DHPLC)
Υγρή χρωματογραφία με υψηλής απόδοσης αποδιάταξη
· Conformation Sensitive Gel Electrophoresis (CSGE).
Ηλεκτροφόρηση σε πηκτή με ευαισθησία διαμόρφωσης
· Universal Heteroduplex Generator(UHG)
Καθολική Γεννήτρια Ετερόδιπλου μορίου
Οι βασικές αρχές αυτών των τεχνικών περιγράφονται στη συνέχεια. [1]
Υβριδισμός με ειδικά ολιγονουκλεοτίδια. (Allele Specific Oligonucleotides)(ASO)
Αυτή η μέθοδος χρησιμοποιείται για να ανιχνεύσει μια ή περισσότερες γνωστές σημειακές μεταλλαγές στον πληθυσμό. Χρησιμοποιεί τους ελέγχους των νουκλεοτιδίων που είναι κοντά σε συγκεκριμένες και ιδιαίτερες ακολουθίες DNA. Αυτή είναι η απλούστερη και η λιγότερο ακριβής προσέγγιση από το να ψάχνουμε για οποιαδήποτε νέα μεταλλαγή στα μέλη μιας οικογένειας. (Εικόνα 1) Το σημαντικότερο μειονέκτημα αυτής της τεχνικής είναι ότι χρησιμοποιεί ενεργό ράδιο και μπορεί να προκαλέσει άλλες μεταλλαγές εάν αυτό είναι παρόν. [2]

Εικόνα 1 [2]
Επίσης το πρωτόκολλο της ASO, στηρίζεται στη διάκριση μεταξύ της διαφοράς δύο μορίων DNA σε μια βάση κατά την υβριδοποίηση. PCR φθορισμού εφαρμόζεται στα τμήματα που λέγονται SNP ακολουθίες. Μετά από αυστηρή υβριδοποίηση και πλύση, μετριέται η ένταση του φθορισμού για κάθε ένα SNP ολιγονουκλεοτίδιο. (Εικόνα 2)

Εικόνα 2 [3]
Πρωτεϊνική δοκιμή αποκοπής (Protein truncation test) (PTT)
Οι περισσότερες BRCA1 και BRCA2 μεταλλαγές έχουν σαν αποτέλεσμα μικρότερη ή περικεκομμένη πρωτεϊνη.Η PTT είναι μια μέθοδος που ανιχνεύει τις μεταλλαγές που οφείλονται στη πρόωρη λήξη της μετάφρασης του mRNA. Ως αποτέλεσμα αυτού, το πρωτεϊνικό προϊόν είναι περικεκομμένο. Η περικεκομμένη πρωτεϊνη θα μπορούσε να προκύψει λόγω μιας μεταλλαγής που μετατοπίζει το πλαίσιο ανάγνωσης, μιας μη κωδικής μεταλλαγής και μιας μεταλλαγής θέσης του ματίσματος. Το πλεονέκτημα της μεθόδου PTT είναι το ότι μπορεί να ανιχνεύσει τις μεταλλαγές πολλών κιλοβάσεων. Εντούτοις, αυτή η μέθοδος δεν μπορεί να ανιχνεύσει τους πολυμορφισμούς, τις σιωπηλές μεταλλαγές και τις παρερμηνεύσιμες μεταλλαγές. [4]
Οι περισσότερες ασθένειες-προκαλούνται σαν αποτέλεσμα μεταλλαγών αποκοπής του πρωτεϊνικού προϊόντος, έτσι η πρωτεϊνική δοκιμή αποκοπής (PTT) χρησιμοποιείται συχνότερα επειδή θα ανιχνεύσει τις αλλαγές που είναι βιολογικά σημαντικές. Είναι μέθοδος που επιλέγεται για τις μεταλλαγές ογκοκατασταλτικών γονιδίων όπου πάνω από το 90-95% των μεταλλαγών είναι λήξη αλυσίδων (BRCA1, BRCA2, APC). Το γονίδιο (καλύτερα να αρχίζουμε με mRNA ως πρότυπο) ενισχύεται από PCR και το προϊόν από την PCR αντίδραση χρησιμοποιείται για να μεταγράψει / μεταφράσει την πρωτεϊ'νη που κωδικοποιείται από το γονίδιο χρησιμοποιόντας ένα in vitro συστήματος μεταφράσεων.(Εικόνα 3) Η συντεθειμένη πρωτεϊνη οργανώνεται από SDS και έναντι της πρωτεϊνης άγριου-τύπου όπου οποιεσδήποτε διαφορές στο μέγεθος μπορούν να απεικονιστού με χρώση στο πήκτωμα με Coomassie μπλε ή ασήμι χρωστική.[5]

Εικόνα 3 [5]
Πολυμορφισμός διαμόρφωσης μονόκλωνης αλυσίδας (Single Strand Conformational Polymorphism)(SSCP)
Αυτή η μέθοδος μπορεί να ανιχνεύσει μεταλλαγές σε βάσεις μονόκλωνης αλυσίδας όπως τις μεταλλαγές μετατόπισης του πλαισίου ανάγνωσης, μη κωδικές μεταλλαγές και κωδικές μεταλλαγές στον καρκίνο του μαστού. Η βασική αρχή αυτής της δοκιμής είναι το ότι το δίκλωνο DNA όταν αποδιατάσεται, παίρνει μια ειδική διαμόρφωση. Αυτή η διαμόρφωση είναι μοναδική και εξαρτάται από την αρχική ακολουθία νουκλεοτιδίων. Αυτή η μέθοδος είναι αρκετά ευαίσθητη για να ανιχνεύσει ακόμη και διαφορά στις μονόκλωνες νουκλεοτιδικές αλυσίδες που καταλαμβάνουν μια διαφορετική διαμόρφωση και όταν υποβάλλονται σε ηλεκτροφόρηση, η παραλλαγή των νουκλεοτιδίων καταλαμβάνει μια διαφορετική θέση. [6]
Μια αλλαγή ενός νουκλεοτιδίου σε μια συγκεκριμένη ακολουθία, όπως φαίνεται στο δίκλωνο DNA δεν μπορεί να διακριθεί με ηλεκτροφόρηση, γιατί οι φυσικές ιδιότητες της διπλής έλικας είναι σχεδόν ίδιες και για τα δύο αλληλόμορφα γονίδια. Αντιθέτως, μετά από την αποδιάταξη, η διπλή έλικα υποβάλλεται σε τρισδιάστατο δίπλωμα και μπορεί να πάρει διαφορετικές διαμορφώσεις σύμφωνα με την αλληλουχία του DNA. Ανάλογα με αυτήν την διπλωμένη δομή που τα μόρια έχουν πάρει, μπορούν να ταξιδεψουν γρηγορότερα ή πιό αργά σε ένα πήκτωμα, ακόμα κι αν ο αριθμός νουκλεοτιδίων είναι ο ίδιος. [7]
Η SSCP ήταν ένας τρόπος να ανακαλυφθούν νέοι ξεχωριστοί πολυμορφισμοί DNA με DNA αλληλούχιση, αλλά αντικαθίσταται τώρα με τεχνικές αλληλούχισης εξ αιτίας της αποδοτικότητας και λόγων ακρίβειας. Επίσης, η SSCP μπορεί να χρησιμοποιηθεί στην γενοτύπηση, στην ανίχνευση ομοζυγωτικών ατόμων από διαφορετικές καταστάσεις αλληλόμορφων, όπως και ετερογυζυγωτικών ατόμων και πρέπει να καταδείξει ο κάθε ένας ευδιάκριτα σχέδια σε ένα πείραμα ηλεκτροφόρησης.[7]
Η SSCP, έχει αποδειχθεί ένα χρήσιμο εργαλείο για την ανίχνευση των SNPs. Έχει χρησιμοποιηθεί επιτυχώς για την ανίχνευση άγνωστων μεταλλαγών και για εξέταση γνωστών μεταλλαγών. Η τεχνική είναι απλή και εφαρμόζεται καλά σε οποιοδήποτε εργαστήριο όπου εκτελείται αλληλούχηση και γενοτύπηση. Στην PCR-SSCP ανάλυση, το ενισχυμένο προϊόν από ένα τμήμα DNA αποδιατάσσεται και εμποδίζεται η επαναδιάταξή του με το συμπληρωματικό του, με διάλυση, κατάψυξη, ή την προσθήκη παραγόντων αποδιάταξης όπως το NaOH. Αυτό αναγκάζει κάθε κλώνο σε επαναδιάταξη από μόνο του σε μία χαρακτηριστική σταθερή διαμόρφωση. Όταν αυτές οι διαμορφώσεις ηλεκτροφορηθούν κάτω από φυσικές συνθήκες (μη αποδιάταξης), η κινητικότητα κάθε κλώνου επηρεάζεται από τη διαμορφωμένη δομή του. Αυτή η δομή εξαρτάται από την ακολουθία των νουκλεοτιδίων των κλώνων (Εικόνα 4 ). Παραλλαγή ακολουθίας με PCR προϊόν, όπως μια σημειακή μεταλλαγή, παράγει μία διαφορετική διαμόρφωση, με συνέπεια μια αλλαγή στην ηλεκτροφορητική κινητικότητα. Με τη σύγκριση της κινητικότητας των κλώνων μεταξύ των δειγμάτων, μπορεί να προσδιοριστεί μια γενωμική παραλλαγή.[8]
 [8]
[8]  [9]
[9]
Εικόνα 4 . SSCP ανάλυση. Κάθε αλληλόμορφο γονίδιο από την PCR ενίσχυση διαμορφώνει μιά μοναδική σταθερή διαμόρφωση ανάλογα με την ακολουθία. μιας απλής αλλαγή νουκλεοτιδίου που αλλάζει τη διπλωμένη δομή κάθε αποδιαταγμένης αλυσίδας, με μια συνακόλουθη αλλαγή στην ηλεκτροφορητική κινητικότητα. [8]
SSCP έχει εφαρμοστεί παραδοσιακά στα πρότυπα πηκτώματα πολυακρυλαμιδίου, με χρήση του p32 με ασημί χρώση, ή με χρώση με βρωμίδιο του εθιδίου. (Εικόνα 5) Μια εναλλακτική λύση είναι η SSCP φθορισμού (Εικόνα 6 ). Η SSCP φθορισμού έχει τρία σημαντικά πλεονεκτήματα σε σχέση με τις μονοχρωματικές μεθόδους στην p32 ή στην ασημί χρώση. Πρώτα, PCR εκκινητές μπορούν να ιχνηθετηθούν με φθορισμό και η ενίσχυση να τρέχει με αυτοματοποιημένη αλληλούχηση, ‘κόβωντας δρόμο’ με την εξάλειψη του μετέπειτα χειρισμού με τα πηκτώματα και την εξάλειψη της χρήσης επικίνδυνων πηγών ακτινοβολίας και τις τοξικές ασημί -χρώσης χημικές ουσίες. Δεύτερον, κάθε ένας PCR εκκινητής ιχνηθετείται με ένα διαφορετικό χρώμα, επιτρέποντας την ανίχνευση λεπτές μετατοπίσεις κινητικότητας ή μετατοπίσεις στις οποίες η μεταλλαγμένη αλυσίδα αναστρέφει τις κινητικότητες με τις αλυσίδες άγριου - τύπου. Το τρίτο σημαντικό πλεονέκτημα είναι ότι ένας έλεγχος μπορεί να είναι ιχνηθετημένος με φθορισμό με ένα φθορίζον τρίτιο και να τρέχει σε κάθε δίαϋλο. Ο έλεγχος μπορεί να γίνει με εμπορικά πρότυπα όπως τα GS350 μεγέθους πρότυπα (PE Biosystems, Foster City, CA, USA), ή για ακόμα μεγαλύτερη ευαισθησία, γνωστά άγριου τύπου - δείγματα με φθορίζον τρίτιο μπορούν να ενισχυθούν με PCR. Ένας εσωτερικός έλεγχος καθιστά πιθανή την ανάπτυξη δειγμάτων. Τα SSCP φθορισμού πηκτώματα αναλύεται με το GeneScan (PE Biosystems) και σημειώνονται αποτελεσματικά με το λογισμικό Genotyper. [8]

Εικόνα 5 PCR-SSCP ανάλυση ανίχνευσης μεταλλαγής. Σε αυτό των 220-bp τμήμα, μιά από C σε T αντικατάσταση στο δείγμα 7 προκαλεί μια αλλαγή στην διαμορφωμένη δομή κάθε κλώνου. Αυτή η αλλαγή οδηγεί σε μιά δραματική μετατόπιση κινητικότητας στη χαμηλότερη ζώνη και μια λεπτή μετατόπιση στην ανώτερη ζώνη [8]

Εικόνα 6 SSCP ανάλυση φθορισμού. Άγριου-τύπου DNA ενισχύεται με HEX-ιχνηθετημένους εκκινητές (παχιές γραμμές) και τρέχει σε κάθε κανάλι ελέγχου. Κανονικά, δείγματα με ετερογυζυγωτική μετάλλαξη και με ομόζυγωτική μετάλλαξη ενισχύονται με TET-ιχνηθετημένο forward εκκινητή (στικτές γραμμές) και FAM - ιχνηθετημένο reverse εκκινητή (συνεχείς γραμμές). Υπό αυτούς τους όρους, μόνο μόνο ο forward κλώνος διαμορφώνεται, όπως υποδεικνύεται από την κινητικότητα της μετατόπισης της πράσινης ζώνης. [8]
Αλληλούχηση νουκλεοτιδίων (Nucleotide sequencing)
Σε αυτήν την μέθοδο, το πλήρες γονίδιο αλληλουχίζεται για να προσδιοριστεί η παρουσία από οποιαδήποτε μεταλλαγή. Μέχρι την στιγμή της πλήρους αλληλούχησης του γονιδίου εξετάζεται σαν μέθοδος χρυσών προτύπων επειδή οποιοσδήποτε τύπος μεταλλαγής μπορεί να προσδιοριστεί. Η αλληλούχηση γίνεται με δύο μεθόδους:
Sanger's διδέοξυ νουκλεοτιδίου μέθοδος
Shotgun μέθοδος [7]
Η αρχή πίσω από αυτήν την τεχνική είναι: οι κλώνοι του DNA μπορούν να χωριστούν μεταξύ τους με αποδιάταξη. Ομοίως, μπορούν επίσης να επαναδιαταχθούν, όταν οι κλώνοι βρεθούν πάλι μαζί, με άλλα λόγια, να επανενωθούν για να διαμορφώσουν ομόδιπλη δομή. Αλλά εάν υπάρχει μια μεταλλαγή στον έναν από τους κλώνους, το συνιστάμενο DNA θα είναι σε ετερόδιπλη κατάσταση.(Εικόνα 8) Μια τέτοια ιδιότητα του DNA μπορεί να χρησιμοποιηθεί καλύτερα για την ανίχνευση των σημειακών μεταλλαγών. Επιπλέον, οι ετερόδιπλοι DNA κλώνοι κινούνται πιο αργά επάνω σε πήκτωμα πολυακρυλαμιδίου όταν συγκρίνεται με το ομόδιπλο DNA.[10]

Εικόνα 8 [11]
Τεχνολογία DNA μικροσυστοιχιών (chips) (DNA microarray technology)
Η σύνδεση των βάσεων ή η συμπληρωματικότητα των βάσεων είναι η αρχή πίσω από αυτή τη νέα τεχνολογία. Οι πιθανές εφαρμογές της microarray τεχνολογίας είναι η σκιαγράφηση της έκφρασης των γονιδίων και ο προσδιορισμός των ακολουθιών των γονιδίων (συμπεριλαμβανομένων των ακολουθιών που αφορούν τις μεταλλαγές). Σε αυτήν την μέθοδο, ένας μεγάλος αριθμός των DNA τμημάτων τοποθετείται σε μια φωτογραφική διαφάνεια γυαλιού. Τα ταμήματα αφήνονται να υβριδώσουν ή να συνδεθούν με ιχνηθετημένο DNA (έλεγχοι), που υβριδιοποιούν το DNA στη φωτογραφική διαφάνεια γυαλιού. Το ποσό της υβριδοποίησης αναλύεται έπειτα σε κάθε σημείο στη φωτογραφική διαφάνεια. Στα γονίδια δίνεται ένα χρώμα όπου τα καλά υβριδοποιημένα χρωματίζονται κόκκινα και τα γονίδια που υβριδοποιούνται λιγότερο είναι χρωματισμένα πράσινα. Η microarray τεχνολογία θα μπορούσε να χρησιμοποιηθεί στον έλεγχο και την ταξινόμηση της ενίσχυσης των χρωμοσωμάτων και της έλλειψης αυτών, στους όγκους, στην ανακάλυψη φαρμάκων και την ανάπτυξή τους. Επίσης η τεχνική εφαρμόζεται πρακτικά στην ανίχνευση των μεταλλαγών και την περαιτέρω έρευνα του μηχανισμού της ανάπτυξης των όγκων. [12]
Δυνατότητα προσδιορισμού δεκάδων, εκατοντάδων ή χιλιάδων αλλαγών στα DNA, RNA, και, εσχάτως,στις πρωτεΐνες, σε 1 δοκιμασία (φυσικά, στο ίδιο δείγμα)
Βασική τεχνολογία
Φωτολιθογραφία μερομποτική
–pl συγκεκριμένων αλληλουχιών (10 άδων, 100 άδων, 1000 άδων) σταθεροποιούνται σε ένα πλακίδιο ή νάυλον μεμβράνη
•Ανάλογα με το μήκος / προέλευση αλληλουχιών
–Oligo-arrays
· Expression
· SNPs
· Sequencing
· Genotyping
–cDNAarrays
· Expression arrays
· CGH arrays
–Protein-arrays
· Πολλαπλά software για την ανάλυση
Σύγκριση μεκλασσικές μοριακές μεθόδους
· Βασική αρχή (και στα arrays): συμπληρωματικότητα βάσεων νουκλεϊνικών οξέων
· Αντί για 1 ελέγχονται 1000άδες στόχοι ταυτόχρονα στο ίδιο δείγμα
· Απαραίτητη η ρομποτική (για την κατασκευή) και η βιοπληροφορική (για την ανάλυση).
Δυνατότητες
Ερευνητικά: ΕΠΑΝΑΣΤΑΣΗ!
· Γονοτυπική ανάλυση και προσδιορισμός υποτύπων καρκίνου που σχετίζονται με απάντηση σε θεραπεία
· Ανάλυση μεταλλάξεων γνωστών γονιδίων που εμπλέκονται σε συγκεκριμένο καρκίνο
· Εντοπισμός νέων γονιδίων – στόχων για ανάπτυξη φαρμάκων
· Εντοπισμός νέων γονιδίων που μπορεί να χρησιμεύσουν για παρακολούθηση / πρόληψη
· Αναγνώριση και αξιολόγηση προφίλ γενετικών αλλαγών για τη χορήγη σησυγκεκριμένης περαιτέρω θεραπείας
Διαγνωστικά
· Χρήση των παραπάνω για την αξιολόγηση της κατάστασης τουασθενούς και την επιλογή θεραπείας.[13]

Εικόνα 9 [14]
Ηλεκτροφόρηση σε πηκτή βαθμιδωτής θερμοκρασίας (Temperature gradient gel electrophoresis) (TGGE)
Είναι μια μορφή ηλεκτροφόρησης που μελετά τη συμπεριφορά των ουσιών κάτω από διαφορετικές θερμοκρασίες. Η TGGE είναι μια από την οικογένεια των ηλεκτροφορητικών μεθόδων για το χωρισμό των νουκλεϊνικών οξέων όπως το DNA ή το RNA που στηρίζεται σις εξαρτώμενες από την θερμοκρασία αλλαγές στη δομή. Η αρχική μέθοδος ήταν η DGGE, που είναι σχεδόν ίδια.(Εικόνα 10) Η DGGE εφευρέθηκε από τον Leonard Lerman, ενώ ήταν καθηγητής στο SUNY Albany. Ενώ ο ίδιος εξοπλισμός μπορεί να χρησιμοποιηθεί για την ανάλυση των πρωτεϊνών, που παραγάγουν παρόμοια στην όψη σχέδια, οι θεμελιώδεις αρχές είναι αρκετά διαφορετικές για τις πρωτεϊ'νες και τα νουκλεϊνικά οξέα (ο T Creighton από το MRC εργαστήριο στην Αγγλία ήταν το πρώτο άτομο που έκανε αυτό). Από τότε οι δύο αυτές μέθοδοι συσχετίζονται γραμμικά,, και είναι από μια θεωρητική σκοπιά, σχεδόν πανομοιότυπες. Κατά συνέπεια, για να καταλάβουμε την TGGE θα ήταν καλύτερα πρώτα να εξετάσουμε τις αρχές της DGGE. Η TGGE αναπτύχθηκε αρχικά από τους Lerman και Andersen με χρήση πιάτο από οξείδιο του βηρυλλίου σαν θερμικό διασκορπιστή (το BeO έχει μια πολύ υψηλή θερμική αγωγιμότητα) και από τον Roger Wartell του Georgia Tech. Extensive η δε εργασία έγινε από την ομάδα Riesner στην

Εικόνα 10 [15]
Για να καταλάβει κανείς την T/DGGE, υπάρχουν δύο θεμελιώδη σημεία. Το πρώτο είναι πώς η δομή του DNA αλλάζει με τη θερμοκρασία, και το δεύτερο είναι πώς αυτές οι αλλαγές στη δομή έχουν επιπτώσεις στη μετακίνηση του DNA μέσω ενός πηκτώματος. Αρχίζουμε με ένα μόριο δίκλωνου DNA με μερικές εκατοντάδες ζεύγη βάσεων σε μήκος. Στη θερμοκρασία δωματίου, παρουσία τουλάχιστον ενός mM από άλας, η δίκλωνη μορφή είναι αρκετά σταθερή, και μπορούμε να εξετάσουμε το μόριο που είναι δύο σειρές στενά τυλιγμένες μεταξύ τους, ενεργά οι δύο άκρες. του DNA είναι αρνητικά φορτισμένες (ανιόν) και στην παρουσία ενός ηλεκτρικού πεδίου, θα κινηθεί προς το θετικό ηλεκτρόδιο. Ένα πήκτωμα είναι ένα μοριακό πλέγμα, με τις τρύπες κατά προσέγγιση με το ίδιο μέγεθος με τη διάμετρο του DNA. Παρουσία του ηλεκτρικού πεδίου, το DNA θα προσπαθήσει να κινηθεί μέσω του πλέγματος, και για ένα δεδομένο σύνολο συνθηκών, η ταχύτητα της μετακίνησης είναι κατά προσέγγιση ανάλογη προς το μήκος του DNA μορίου - αυτή είναι η βάση για τον από το μέγεθος εξαρτώμενο χωρισμό στην τυποποιημένη ηλεκτροφόρηση. Καθώς κάποιος αυξάνει τη θερμοκρασία, οι δύο κλώνοι του DNA αρχίζουν να χωρίζουν και αυτό λειώνει. Σε κάποια υψηλή θερμοκρασία, οι δύο κλώνοι θα χωρίσουν εντελώς. Εντούτοις, σε κάποια ενδιάμεση θερμοκρασία, οι δύο κλώνοι θα χωριστούν εν μέρει, με μέρος του μορίου να είναι ακόμη δίκλωνο και ένα άλλο μέρος μονόκλωνο. Κάποιος θα μπορούσε να το κάνει αυτό από το ένα τέλος, για να καταστήσει μία Υ διαμορφωμένη δομή με 3 άκρα, και από τις δύο άκρες για να κάνουν μια δομή με 4 άκρες, ή στη μέση για να κάνει μια φυσαλίδα. Αυτό που κάνει την D/TGGE επωφελή είναι ότι η κινητικότητα του DNA μορίου μέσω του πηκτώματος μειώνεται δραστικά όταν διαμορφώνονται σε αυτό μερικώς λειωμένες δομές, και το σημαντικότερο, η ακριβής θερμοκρασία στο οποίο αυτό εμφανίζεται εξαρτάται από την αλληλουχία. Κατά συνέπεια η D/TGGE προσφέρει μια μέθοδο εξαρτώμενη από την ακολουθία, ανεξαρτήτως μεγέθους για το χωρισμό των μορίων DNA.
Η DGGE δουλεύει με την εφαρμογή ενός μικρού δείγματος DNA (ή RNA) σε ένα πήκτωμα ηλεκτροφόρησης που περιέχει έναν παράγοντα αποδιάταξης. Οι ερευνητές έχουν διαπιστώσει ότι ορισμένα πηκτώματα αποδιάταξης είναι ικανός να προκαλέσουν «λειώσιμο» του DNA σε ποικίλες καταστάσεις. Ως αποτέλεσμα αυτής της τήξης, το DNA μετακινείται μέσω του πηκτώματος και μπορεί να αναλυθεί στα απλά συστατικά του, ακόμη και σε εκείνα τα τόσο μικρά όπως των 200-700 ζευγών βάσεων.
Περαιτέρω εξηγώντας πώς αυτή η τεχνική λειτουργεί σημειώνουμε κάτι που είναι μοναδικό για την DGGE τεχνική που είναι ότι καθώς το DNA υποβάλλεται όλο και περισσότερο extreme συνθήκες αποδιάταξης, τα τμήματα των κλώνων ξεδιπλώνουν εντελώς σε ένα απλό κλώνο. Αυτή η διαδικασία είναι μοναδική επειδή έρχεται σε αντίθεση με το πώς το DNA αποδιατάσσεται in vivo. Λόγω αυτής της ποιοτικής διάκρισης του DNA όταν τοποθετείται σε πηκτώμα αποδιάταξης, είναι δυνατό για τους ερευνητές να διακρίνουν τις διαφορές στις ακολουθίες του DNA ή στις μεταλλαγές των διάφορων γονιδίων.[16]
Η (DGGE) είναι ένα εργαλείο που αναπτύχθηκε για να αναλύσει το DNA. Το DNA είναι ένα μόριο που είναι μια διπλή έλικα από δύο συμπληρωματικούς κλώνους που μοιάζει με ένα φερμουάρ ιματισμού που έχει στριφτεί κατά μήκος του άξονά του. Η DGGE χρησιμοποιείται για να ανιχνεύσει τις αλλαγές (μεταλλαγές) στο γενετικό κώδικα μέσα σε ένα δείγμα, και μπορεί να ανιχνεύσει τόσο μικρές διαφορές, όσο ένα ζεύγος βάσεων μεταξύ των κλώνων του DNA.
Πώς η διαδικασία βρίσκει τις μεταλλαγές
Η αρχή αυτής της τεχνικής είναι να χωρίσει τους DNA κλώνους, με βάση την πραγματική σύνθεση των βάσεων, ή την αναλογία των GC προς AT ζευγών βάσεων που συγκροτούν ένα ξεχωριστό τμήμα του DNA. Αυτό ολοκληρώνεται με την έκθεση του DNA σε κλίση αποδιατακτικού με ανυψωμένες θερμοκρασίες μέσα στο πήκτωμα πολυακρυλαμίδιο.(Εικόνα 7)

Εικόνα 7 [17]
Καθώς το δείγμα DNA προχωρεί μέσω του πηκτώματος, από μία χαμηλή αποδιατακτική συγκέντρωση σε μια μεγαλύτερη, αρχίζει να λειώνει σε διάφορα σημεία. Αυτό είναι συγγενές με την DNA αποσυμπίεση (“unzipping.”) Υψηλότερο GC περιεχόμενο από το δείγμα, καθιστά δυσκολώτερο το λειώσιμο. Κατά συνέπεια, το δείγμα DNA είναι σε θέση να προχωρήσει περαιτέρω στο πήκτωμα πρίν σταματήσει. Δείγματα με χαμηλότερο GC περιεχόμενο λειώνουν γρηγορότερα συγκριτικά. Επομένως, προχωρούν πιό αργά μέσα στο πήκτωμα, χωρίζοντας κατά συνέπεια από την άλλη την γρηγορότερα κινούμενη αλυσίδα του DNA.[18]
Είναι σαφές ότι η μέθοδος TGGE χρησιμοποιεί τη θερμότητα ως αρχικό μηχανισμό για το ξετίληγμα και την αποδιάταξη του DNA. Αυτό που δεν είναι προφανές εντούτοις, είναι το πως αυτή η διαδικασία εμφανίζεται και το πώς το DNA μπορεί να αναλυθεί χρησιμοποιόντας αυτή την τεχνική. Αυτό που είναι ίσως το πιό ενδιαφέρον κατά την εξέταση της διαδικασίας της αποδιάταξης με την TGGE μέθοδο είναι ότι εμφανίζεται σε τέτοιες συστηματικές διαδικασίες, που είναι δυνατόν να αναδημιουργήσει τα τμήματα μόλις αυτά χωριστούν. Η εργασία με PCR ηλεκτροφόρηση των τμημάτων ξεκινά με μόρια διπλής έλικας. Σε μια ορισμένη θερμοκρασία, το DNA αρχίζει να λειώνει, με κατάληξη σε μια δομή ομοιάζουσα με φούρκα. Σε αυτήν την διαμόρφωση η απομάκρυνση επιβραδύνεται συγκρινόμενη με το εντελώς δίκλωνο τμήμα του DNA. Έκτοτε η θερμοκρασία τήξης εξαρτάται σημαντικά από την ακολουθία των βάσεων, των τμημάτων του DNA του ίδιου μεγέθους αλλά οι διαφορετικές ακολουθίες μπορούν να χωριστούν. Κατά συνέπεια η TGGE όχι μόνο χωρίζει τα μόρια, αλλά δίνει τις πρόσθετες πληροφορίες σχετικά με την τήξη, τη συμπεριφορά και την σταθερότητα.[16] (Εικόνα 11)


Εικόνα 11 : Αρχές της TGGE, και προφίλ της μετακίνησης σε TTGE των τεμαχίων του DNA σε βακτήρια. [19]
Οι πληροφορίες αυτές εξυπηρετούν τη θεμελίωση για την κατανόηση του θεωρητικού πλαίσιου πίσω από την TGGE. Όταν πάμε να δημιουργήσουμε μιά TGGE, είναι σαφές ότι η μεθοδολογία υιοθετείται σχεδόν ξεκάθαρα σαν αρχή που κρύβεται κάτω από την αυτήν τεχνική. Συνοψίζοντας, τα βήματα που απαιτούνται σχετικά με την παραγωγή δειγμάτων της TGGE, είναι:
· Ρίψη των πηκτωμάτων - Αυτό το βήμα απαιτεί ένα άτομο για να προετοιμάσει τους δοκιμαστικούς σωλήνες για τη μηχανή, να προετοιμάσει το πήκτωμα για τη μηχανή και να χύσει το πήκτωμα για την ηλεκτροφόρηση. Από όλα αυτά τα βήματα, προετοιμάζοντας το πήκτωμα φαίνεται να τίθεται η σημαντικότερη πρόκληση για τον ερευνητή. Επειδή ένα ρυθμηστικό σύστημα πρέπει να επιλεχτεί, είναι σημαντικό το σύστημα να παραμένει σταθερό μέσα το πλαίσιο της αυξανόμενης θερμοκρασίας. Κατά συνέπεια, χρησιμοποιείται χαρακτηριστικά η ουρία για την προετοιμασία των πηκτωμάτων. Εντούτοις, οι ερευνητές πρέπει να γνωρίζουν ότι το ποσό της ουρίας που χρησιμοποιείται θα ασκήσει επίδραση στη γενική θερμοκρασία που απαιτείται για να χωρίσει το DNA. Ανάλογα ποιος τύπος TGGE είναι να οργανωθεί, κάθετος ή παράλληλος, ποικίλουν τα ποσά των δειγμάτων που είναι ανάγκη να προετοιμαστούν και να φορτωθούν. Ένα μεγαλύτερο ποσό ενός δείγματος χρησιμοποιείται με τον κάθετο, ενώ ένα μικρότερο ποσό πολλών δειγμάτων χρησιμοποιείται με τον παράλληλο τύπο TGGE.
· Ηλεκτροφόρηση - Αυτό το βήμα είναι αυτεξήγητο. Στο πήκτωμα φορτώνεται, το δείγμα τοποθετείται στη θέση χορήγησης των πηκτωμάτων στον τύπο του πηκτώματος που οργανώνεται —π.χ.. παράλληλο ή κάθετο — η τάση ρυθμίζεται και το δείγμα μπορεί να αφεθεί να τρέξει.
· Κηλίδωση – Μόλις οργανωθεί το πήκτωμα, για να κρατηθούν τα αποτελέσματα σταθερά και περαιτέρω για να είναι σε θέση να διαβαστούν, το πήκτωμα πρέπει να κηλιδωθεί. Ενώ υπάρχει ένας αριθμός από τους κηλίδες που μπορούν να χρησιμοποιηθούν για αυτόν το λόγο, η ασημί χρώστική έχει αποδειχθεί το αποτελεσματικότερο εργαλείο.
· Εκλεκτική προσρόφηση του DNA – Σε αυτό το βήμα το DNA μπορεί να διαχωριστεί με εκχύλιση από την ασημί χρωστική για την περαιτέρω ανάλυση μέσω της PCR ενίσχυσης.[16]
Εξέταση της εφαρμογής αυτής της τεχνολογίας μέσα στο ευρύτερο πλαίσιο της ιατρικής επιστήμης, είναι σαφές ότι η TGGE έχει ένα ευρύ πεδίο χρησιμότητας στην επιστημονική έρευνα. Για να επεξηγηθεί αυτό το σημείο, ένα μόνο χρειάζεται για να εξετάσει την τρέχουσα έρευνα στο θέμα. Με την εξέταση του πώς η TGGE εφαρμόζεται στην πρακτική έρευνα είναι δυνατό να γίνουν κατανοητά τα οφέλη αυτής της τεχνολογίας για την πρόοδο της επιστήμης και της ιατρικής φροντίδας.[16]
Σύμφωνα με μια πρόσφατη έρευνα των Wong, Liang, Kwon, Bai, Alper and Gropman, η TGGE μπορεί να χρησιμοποιηθεί για να εξετάσει το μιτοχονδριακό DNA από ένα άτομο. Η TGGE χρησιμοποιήθηκε για να καθορίσει δύο ασυνήθιστες μεταλλαγές στο μιτοχονδριακό γονιδίωμα : "Μιά 21 χρονών γυναίκα που υπήρχαν υποψίες για μιτοχονδριακές κυτταροπαθείς, αλλά όχι για κοινές μιτοχονδριακές σημειακές μεταλλαγές του DNA (mtDNA) και διαγραφές, μελετήθηκε για άγνωστες μεταλλαγές σε ολόκληρο το μιτοχονδριακό γονιδίωμα με TGGE. Μέσω της TGGE έγινε προφανές ότι αυτός ο ασθενής έχει μια μεταλλαγή, εντούτοις δεν ανέπτυξε ακόμα κυστική ίνωση. Αυτές οι πληροφορίες όχι μόνο παρέχουν μερικές προγνώσεις στην πρόοδο της κυστικής ίνωσης στα άτομα που έχουν αυτές τις μεταλλαγές γονιδίων, αλλά και καταδεικνύουν ότι η κυστική ίνωση είναι μια ασθένεια ολόκληρου του μιτοχονδριακού γονιδιώματος.[20]
Ο Lohr και οι συνάδελφοί του (2001) αναφέρουν σε μια περιεκτική μελέτη για τα παγκρεατικά υγρά από άτομα χωρίς παγκρεατικό καρκίνωμα, ότι p53 μεταλλαγές, θα μπορούσε να βρεθεί στα παγκρεατικά υγρά σε ένα μικρό ποσοστό των συμμετεχόντων. Επειδή οι μεταλλαγές p53 συναντώνται εκτενώς στα παγκρεατικά καρκινώματα, οι ερευνητές για αυτήν την έρευνα προσπαθούσαν για να καθορίσουν εάν η μεταλλαγή από μόνη της μπορεί να συνδεθεί με την ανάπτυξη του καρκίνου στο πάγκρεας. Ο Lohr βρήκε p53 μεταλλαγές μέσω TGGE σε μερικές μόνο περιπτώσεις. Κατά συνέπεια, οι ερευνητές καταλήγουν στο συμπέρασμα ότι η p53 μεταλλαγή μπορεί να μην είναι ο μόνος δείκτης της παγκρεατικής ογκογένεσης καρκινωμάτων.[20]
Ενώ διάφορες μελέτες έχουν χρησιμοποιήσει TGGE με σκοπό τη μελέτη εσωτερικοί μηχανισμοί της μεταλλαγής και των διαφορών στο DNA στο ανθρώπινο σώμα, η τεχνική επίσης εφαρμόζεται ευρέως στον τομέα της βιολογίας και της μικροβιολογίας κυττάρων. Οι Cheung και Kinkle (2001) εξέτασαν την απομόνωση του Mycobacterium sp. μέσω της διαδικασίας της TGGE. Σύμφωνα με τους ερευνητές, λίγα είναι γνωστά σχετικά με τους Mycobacterium πληθυσμούς επειδή το είδος αυξάνεται και πεθαίνει τόσο γρήγορα. Κατά συνέπεια, οι ερευνητές απομόνωσαν το έναν βακτηριδιακό κλώνο και τα συνέκριναν με τέσσερα μολυσμένα δείγματα για να σκιαγραφήσουν τις διαφορές στη μοριακή δομή. Οι ερευνητές ήταν σε θέση να δείξουν ότι εμβολιασμός με ορισμένες χημικές ουσίες που περιορίζουν εδαφολογικά το Mycobacterium θα μπορούσε σημαντικά να βελτιώσει την υποβάθμιση του χώματος λόγω του βακτηριδίου.[20]
Υγρή χρωματογραφία με υψηλής απόδοσης αποδιάταξη (DENATURING HIGH-PERFORMANCE LIQUID CHROMATOGRAPHY) (DHPLC)
Η DHPLC προσδιορίζει τις μεταλλαγές και τους πολυμορφισμούς με βάση την ανίχνευση του σχηματισμού ετερόδιπλων μορίων μεταξύ κακώς συνδυασμένων νουκλεοτιδίων στην διπλή έλικα κατά την PCR ενίσχυση του DNA. Παραλλαγή ακολουθίας δημιουργεί μεικτό πληθυσμό ετερόδιπλων μορίων και ομόδιπλων κατά τη διάρκεια ανασυνδιασμού του άγριου - τύπου και της DNA μετάλλαξης (Εικόνα 12). Όταν αυτός ο μικτός πληθυσμός αναλύεται με HPLC κάτω από θερμοκρασίες μερικής αποδιάταξης, τα ετερόδιπλα μόρια διαχωρίζονται με εκχύλιση από τη στήλη νωρίτερα από τα ομόδιπλα εξαιτίας της μειωμένης θερμοκρασία τήξης τους. Η ανάλυση μπορεί να εκτελεσθεί σε δείγματα ενός ατόμου για να προσδιοριστεί η ετεροζυγωτία, (Εικόνα 13) ή σε μικτά δείγματα για να προσδιοριστεί η παραλλαγή των νουκλεοτιδίων μεταξύ των ατόμων.[21]

Εικόνα 12 . Σχηματισμός ετερόδιπλων μορίων. Άγριου-τύπου (Wt) και μετάλλαξης (Mt) Τα PCR προϊόντα θερμαίνονται για να αποδιαταχθεί κάθε ένας κλώνος και έπειτα αφήνονται να ψυχθούν αργά. Το αποτέλεσμα είναι μικτός πληθυσμός από τα αρχικά ομόδιπλα και ετερόδιπλα μόρια που περιέχουν κακώς συνδυασμένες βάσεις.[21]

Εικόνα 13. Προσδιορισμός ετεροζυγωτίας. Το PCR προϊόν από έναν ετεροζυγώτη φυσικά διαμορφώνει τα ετερόδιπλα μόρια εξαιτίας της παραλλαγής της ακολουθίας σε κάθε αλληλόμορφο γονίδιο. Αυτή η ετεροζυγωτία προσδιορίζεται εύκολα με DHPLC όταν αναλύεται κάτω από θερμοκρασίες μερικής αποδιάταξης. (Συνθήκες: eluent A:
DHPLC είναι μέθοδος υγρής χρωματογραφίας ιοντικού-ζεύγους αντίστροφης φάσης και υψηλής απόδοσης (IP-RP-HPLC). Υπάρχουν δύο στήλες στην αγορά για την εκτέλεση ανάλυσης τμημάτων: η DNASep στήλη και η Zorbax Eclipse dsDNA ανάλυσης στήλη. Η συσκευασία Eclipse στήλης αποτελείται από σφαιρικά μόρια πυριτίου (Zorbax Rx-SIL type B), πυκνά βαλμένα σε στρώσεις με ένα αλειφατικό organosilane στατικής φάσης. Η DNASep στήλη έχει μια στατική φάση αποτελούμενη από πολυστυρόλιο - divinylbenzene copolymer. Ο παράγοντας της ιονικής-ένωσης που χρησιμοποιείται με καθεμία στήλη είναι οξικό άλας triethylammonium (TEAA), που μεσολαβεί συνδεόμενο με το DNA στη στατική φάση. Ακετονιτρίλιο χρησιμοποιείται ως οργανικός παράγοντας για να επιτύχει τον επόμενο χωρισμό του DNA από τη στήλη.
Η DHPLC απαιτεί έναν βιοσυμβατό HPLC σύστημα άντλησης με κλίση με μια πορεία ροής αποτελούμενη από PEEK σωλήνωση, συναρμολογήσεις, και φίλτρα; έναν ανιχνευτή υπεριώδους ακτινοβολίας (UV) ικανό να μετρά από 254 έως 260 nm; και ένας φούρνο στηλών ικανό για τη διατήρηση των θερμοκρασιών από
Για την ανάλυση ετερόδιπλων μορίων με CSGE, τα προϊόντα της PCR ηλεκτροφορούνται σε ένα 1mm λεπτό πήκτωμα με 37 καλά χτένια (FMC) προετοιμασμένα με 10 ή 15% πολυακρυλαμίδιο, 99:1 με αναλογία σε ακρυλαμίδιο (Intermountain Scientific, Kaysville, UT) σε 1,4-bis(acryloyl) πιπεραζίνη (Fluka), 10% γλυκόλη αιθυλενίου (Sigma), 15% φορμαμίδιο (GIBCO), 0.1% persulfate αμμωνίου (U.S. Biochemicals), και 0.07% N,N,N![]() ,N
,N![]() -tetramethylethylenediamine (Sigma) σε 0.5× TTE ρυθμιστικό διάλυμα (44mM Tris/14.5 mM Taurine/0.1 mM EDTA ρυθμιστικό διάλυμα, pH 9.0). Είναι σημαντικό να μην αποστειρωθεί το ρυθμιστικό διάλυμα TTE για να λάβει το βέλτιστο διαχωρισμό σε ετερόδιπλα μόρια και ομόδιπλα. Ο βέλτιστος χρόνος πολυμερισμού είναι περίπου 1 ώρα. Τα προϊόντα της PCR που περιείχαν ετερόδιπλα μόρια αναμίχθηκαν με 3 µl από 10× φορτωμένο stock ρυθμιστικό διάλυμα (10× stock διάλυμα με 30% γλυκερίνη /0.25% bromphenol μπλέ/0.25% cyanol ξυλολίων FF). Τα δείγματα χωρίστηκαν με ηλεκτροφόρηση σε πρότυπες DNA συσκευές αλληλούχισης πηκτωμάτων με 37.5×45-cm γυάλινα πιάτα με χρήση 0.5× TTE σαν ηλεκτρόδιο ρυθμιστικό διάλυμα. Χαρακτηριστικά, μια χτένα με 37 διαδρόμους χρησιμοποιήθηκε και πάνω από 5 PCR προϊόντα διαφορετικών μεγεθών αναμίχθηκαν και φορτώθηκαν σε κάθε διάδρομο. Το πήκτωμα προ – ηλεκτροφορήθηκε για 15 min, και τα δείγματα διαχωρίστηκαν στη θερμοκρασία δωματίου κατά τη διάρκεια του τρεξίματος με 40 W και 6 hr για 10% πηκτώματα, ή 40 W και 8,5 ώρες για 15% πηκτώματα. Μετά από την ηλεκτροφόρηση, το πήκτωμα χρωματίστηκε στο γυάλινο πιάτο με 1 µg/ml βρωμίδιο του εθιδίου για 10 min ακολουθούμενο με ξέπλυμα στο νερό. Ένας φορητός UV πυρσός χρησιμοποιήθηκε για να απεικονίσει τις ζώνες. Το σχετικό τμήμα από το πήκτωμα κόπηκε, και μεταφέρθηκε σε ένα κομμάτι χαρτί αποτύπωσης, και έπειτα απελευθερώθηκε από το χαρτί επάνω στην επιφάνεια ένός διαεικονογράφου με το ξέπλυμα στο νερό. Το πήκτωμα φωτογραφίστηκε για την αποτύπωση των πηκτωμάτων.(Εικόνα 14) [22]
-tetramethylethylenediamine (Sigma) σε 0.5× TTE ρυθμιστικό διάλυμα (44mM Tris/14.5 mM Taurine/0.1 mM EDTA ρυθμιστικό διάλυμα, pH 9.0). Είναι σημαντικό να μην αποστειρωθεί το ρυθμιστικό διάλυμα TTE για να λάβει το βέλτιστο διαχωρισμό σε ετερόδιπλα μόρια και ομόδιπλα. Ο βέλτιστος χρόνος πολυμερισμού είναι περίπου 1 ώρα. Τα προϊόντα της PCR που περιείχαν ετερόδιπλα μόρια αναμίχθηκαν με 3 µl από 10× φορτωμένο stock ρυθμιστικό διάλυμα (10× stock διάλυμα με 30% γλυκερίνη /0.25% bromphenol μπλέ/0.25% cyanol ξυλολίων FF). Τα δείγματα χωρίστηκαν με ηλεκτροφόρηση σε πρότυπες DNA συσκευές αλληλούχισης πηκτωμάτων με 37.5×45-cm γυάλινα πιάτα με χρήση 0.5× TTE σαν ηλεκτρόδιο ρυθμιστικό διάλυμα. Χαρακτηριστικά, μια χτένα με 37 διαδρόμους χρησιμοποιήθηκε και πάνω από 5 PCR προϊόντα διαφορετικών μεγεθών αναμίχθηκαν και φορτώθηκαν σε κάθε διάδρομο. Το πήκτωμα προ – ηλεκτροφορήθηκε για 15 min, και τα δείγματα διαχωρίστηκαν στη θερμοκρασία δωματίου κατά τη διάρκεια του τρεξίματος με 40 W και 6 hr για 10% πηκτώματα, ή 40 W και 8,5 ώρες για 15% πηκτώματα. Μετά από την ηλεκτροφόρηση, το πήκτωμα χρωματίστηκε στο γυάλινο πιάτο με 1 µg/ml βρωμίδιο του εθιδίου για 10 min ακολουθούμενο με ξέπλυμα στο νερό. Ένας φορητός UV πυρσός χρησιμοποιήθηκε για να απεικονίσει τις ζώνες. Το σχετικό τμήμα από το πήκτωμα κόπηκε, και μεταφέρθηκε σε ένα κομμάτι χαρτί αποτύπωσης, και έπειτα απελευθερώθηκε από το χαρτί επάνω στην επιφάνεια ένός διαεικονογράφου με το ξέπλυμα στο νερό. Το πήκτωμα φωτογραφίστηκε για την αποτύπωση των πηκτωμάτων.(Εικόνα 14) [22]

Εικόνα 14 Η επίδραση του μεγέθους των προϊόντων στην ανίχνευση των παραλλαγών ακολουθίας με CSGE. (Ανώτερος) CSGE ανάλυση σε ένα 755-bp PCR προϊόν που περιέχει τις ακολουθίες για τα εξόνια 25 και 26 του γονιδίου COL1A1. Κανένα ετερόδιπλο μόριο δεν ανιχνεύεται. (Χαμηλότερος) CSGE ανάλυση από ένα 276-bp PCR προϊόν που περιέχει τις ακολουθίες για το εξόνιο 25 του ίδιου γονιδίου. DNA από τα ίδια πέντε άτομα αναλύθηκαν και στις δύο περιπτώσεις. Ετερόδιπλα μόρια ανιχνεύθηκαν στα δείγματα 3 και 5. [22]

Εικόνα 15 CSGE μετατοπίσεις στις 13 πρώτες ανιχνευμένες μεταλλάξεις στα σπερματοζωάρια. a, Οικογένεια TT14 και έλεγχος για το εξόνιο 10. b, Οικογένειες TT16, TT10 and TT11 και ένας έλεγχος για το εξόνιο 13. c, Οικογένεια TT15 και έλεγχοι για το εξόνιο 16. d, Οικογένειες TT4 και TT3 επιπλέον έλεγχοι για το εξόνιο 17. e, Οικογένειες TT2 και TT1 επιπλέον έλεγχοι για το εξόνιο
Καθολική Γεννήτρια Ετερόδιπλου μορίου (Universal Heteroduplex Generator)(UHG)
Υπόβαθρο: Η ανάλυση ετερόδιπλων μορίων σε πλάκες πηκτωμάτων (HDA), είναι μια δημοφιλής μέθοδος ανίχνευσης για τις γενετικές μεταλλαγές, που χρησιμοποιεί DNA τμήματα χαρακτηριστικά παραγμένα με PCR για να δημιουργήσει τα ομο - και ετερόδιπλα μόρια με τις διαμορφωτικές διαφορές και από τις αλληλουχίες-εξαρτώμενα ηλεκτροφορητικά προφίλ. Χρήση (UHG) ενισχύει τις παραλλαγές που προκαλούνται από αντικαταστάσεις απλών-βάσεων.
Μέθοδοι: Η HDA-UHG σε πλάκες πηκτώματος τροποποιήθηκε σε μιά αποδοτική μέθοδο. Η επίδραση της χρώσης των χρωστικών ουσιών TOPRO5 και YOPRO1 στην ανάλυση των ετερόδιπλων μορίων μελετήθηκε, καθώς επίσης και μέθοδοι ανίχνευσης της απορροφητικότητας στο φθορισμό από το υπεριώδες με λέιζερ (LIF). Επιπλέον, τα εμπλεκόμενα πολυμερή υδροξυαιθυλικής κυτταρίνης, μεθυλικής κυτταρίνης, και γραμμικού πολυακριλαμιδίου αξιολογήθηκαν χωριστά.
Αποτελέσματα: Αυτή η δοκιμή ήταν σε θέση να ανιχνεύσει την παρουσία μυκητοβακτηρίου tuberculosis και την ευαισθησία στο rifampin άμεσα από τα κλινικά δείγματα σε εντυπωσιακά μειωμένο χρόνο ανάλυσης (30 min vs 2.5 h). Βελτιστοποιημένες συνθήκες που συμπεριλαμβάνουν 0.3% μεθυλική κυτταρίνη σαν μήτρα χωρισμού, κατ’ευθείαν χρώση με χρήση 1 µmol/L YOPRO1, και LIF ανίχνευση για την ποσοτική και αναπαραγώγιμη ανάλυση από απλές αντικαταστάσεις βάσεων σε rifampin αντίσταση - καθορισμό περιοχής από rpoB που δείχνει ανθεκτικό φαινότυπο στο rifampin- στο M. tuberculosis.(Εικόνα 16 )
Συμπεράσματα: Τριχοειδής ηλεκτροφόρηση, συνδυασμένη με HDA-UHG τεχνική, μπορει να είναι σημαντικά για γρήγορη και αποδοτική κλινική διάγνωση του ανθεκτικού στο rifampin – στελέχους tuberculosis. [24]

Εικόνα 16 HDA. Σχηματική αναπαράσταση PCR-HDA δοκιμής για την ανίχνευση της ευαισθησίας σε rifampin του Mtb, με παρουσίαση των χαρακτηριστικών αποτελεσμάτων από την ανάλυση ηλεκτροφόρησης πλακών πηκτωμάτων. S, σε rifampin- ευαίσθητος γονότυπος, R, σε rifampin- ανθεκτικός γονότυπος. [24]
In situ υβριδισμός
Mια μοναδική μορφολογική μέθοδος μοριακής βιολογίας με πολύτιμες κλινικές εφαρμογές
Eισαγωγή
O in situ υβριδισμός, ο οποίος εφαρμόσθηκε για πρώτη φορά το 1969, αποτελεί πολύτιμη μέθοδο μοριακής βιολογίας για τον παθολoγοανατόμο, επειδή επιτρέπει τη μορφολογική εντόπιση της γενετικής πληροφορίας. Eνώ οι κλασσικές τεχνικές της μοριακής βιολογίας, όπως η Southern blot και η αλυσιδωτή αντίδραση πολυμεράσης (Polymerase Chain Reaction, PCR), πιστοποιούν απλώς την παρουσία μιας αλληλουχίας DNA ή RNA, ο in situ υβριδισμός προσδιορίζει επιπλέον σε ποιά και πόσα κύτταρα υπάρχει η αλληλουχία αυτή και σε ποιό υποκυτταρικό διαμέρισμα (κυτταρόπλασμα, πυρήνας) εντοπίζεται. Eπιπρόσθετα, μας πληροφορεί εάν η παρουσία της αλληλουχίας αυτής σχετίζεται με συγκεκριμένες ανωμαλίες σε κυτταρικό και ιστικό επίπεδο. H τεχνική του in situ υβριδισμού βασίζεται στη θεμελιώδη ιδιότητα των νουκλεϊκών οξέων να σχηματίζουν σύμφωνα με το νόμο της συμπληρωματικότητας των βάσεων σταθερά διμερή που ονομάζονται υβρίδια. Tα τελευταία μπορεί να αποτελούνται από δύο αλύσους DNA ή συνδυασμό αλύσων RNA-DNA και RNA-RNA. O in situ υβριδισμός μπορεί να εφαρμοσθεί σε όλα τα είδη υλικού, όπως τα κυτταρικά επιχρίσματα και τις τομές παραφίνης και κρυοστάτη. [25]
Tα βασικά εργαλεία της τεχνικής ονομάζονται δείκτες (probes) και αποτελούν κατάλληλα σημασμένες δίκλωνες (DNA probes) ή μονόκλωνες (RNA probes) αλληλουχίες νουκλεοτιδίων, οι οποίες μπορούν να συνδεθούν σύμφωνα με την αρχή της συμπληρωματικότητας των βάσεων και επομένως, να ανιχνεύσουν την αναζητούμενη αλληλουχία. H επιλογή του κατάλληλου δείκτη εξαρτάται από το είδος της εφαρμογής. Aν αυτή αφορά την αναζήτηση DNA (όπως ιοί DNA, αλληλουχίες γονιδίων ή χρωματοσωμάτων) προτιμάται η χρήση δίκλωνων δεικτών DNA. Aν η εφαρμογή αφορά την ανίχνευση RNA τότε προτιμάται η χρήση δεικτών RNA (riboprobes), δεδομένου ότι τα προκύπτοντα υβρίδια RNA-RNA είναι πιο σταθερά από τα υβρίδια DNA-RNA. Eναλλακτικά, για την ανίχνευση RNA σε αφθονία μπορούν να χρησιμοποιηθούν ολιγονουκλεοτίδια, μονήρη ή μίγμα περισσότερων του ενός. Oι δείκτες δίκλωνου DNA έχουν μήκος 100-400 βάσεις συνήθως, παράγονται με κλωνοποίηση σε πλασμίδια βακτηριδίων και σημαίνονται με την τεχνική nick translation. Για την παραγωγή και σήμανση των δεικτών RNA χρησιμοποιείται κυρίως η τεχνική in vitro transcription με τη βοήθεια κατάλληλα σημασμένων νουκλεοτιδίων. Tέλος, τα ολιγονουκλεοτίδια έχουν μήκος 18-30 βάσεις, συντίθενται χημικά σε ειδική συσκευή και σημαίνονται στο 3 άκρο με τη βοήθεια του ενζύμου τελική τρανσφεράση. H σήμανση των δεικτών γινόταν αρχικά με ραδιοϊσότοπα (κυρίως 35S), αλλά η χρήση αυτών έχει πλέον περιορισθεί και χρησιμοποιούνται μόνο σε ανίχνευση RNA, εφ’ όσον δεν είναι επιτυχής η χρήση μη-ισοτοπικών δεικτών. Στα κλινικά εργαστήρια χρησιμοποιούνται σχεδόν αποκλειστικά οι μη-ισοτοπικοί δείκτες, λόγω των κινδύνων και της κακής ποιότητας του σήματος των ραδιενεργών δεικτών. H σήμανση των μη-ισοτοπικών δεικτών γίνεται συνήθως με βιοτίνη, διγοξυγενίνη ή φθορίζουσες χρωστικές. H παρουσία φθορίζουσας χρωστικής, είτε απ’ ευθείας στο δείκτη είτε σε κάποιο αντιδραστήριο από τα επακόλουθα στάδια ανίχνευσης της αντίδρασης υβριδισμού, επιτρέπει την εφαρμογή της φθορίζουσας παραλλαγής της τεχνικής του in situ υβριδισμού, διεθνώς γνωστής ως Fluorescent in situ Hybridization-FISH. [25]

Eικόνα 17. Bασικά στάδια in situ υβριδισμού [25]
Προετοιμασία του δείγματος
Aυτή περιλαμβάνει την επεξεργασία των κυττάρων του δείγματος με πρωτεολυτικά ένζυμα (πεψίνη ή πρωτεϊνάση K) ή μικροκύματα, ώστε να επιτευχθεί η διαπερατότητα κυτταροπλάσματος και πυρήνα στο δείκτη και η αλληλεπίδραση του με τις αναζητούμενες αλληλουχίες DNA ή RNA (Eικόνα 17). H πρωτεόλυση είναι απολύτως απαραίτητη, όταν η μονιμοποίηση του δείγματος έχει γίνει σε ουδέτερη φορμόλη, ενώ δεν απαιτείται σε μονιμοποίηση με αλκοολούχα μονιμοποιητικά. Aποδιάταξη και υβριδισμός. Στο στάδιο αυτό επιτελείται απομάκρυνση των δύο αλύσων DNA του δείκτη και κυττάρου, ώστε να επιτευχθεί ακολούθως η σύνδεση τους στο κατάλληλο χημικό και θερμικό περιβάλλον. Aνίχνευση αντίδρασης υβριδισμού. Mετά από τη μεθυβριδική έκπλυση των δειγμάτων ακολουθεί η ανίχνευση των σχηματισθέντων υβριδίων που γίνεται, ανάλογα με τη σήμανση του δείκτη, με αυτοραδιογραφία, ιστοχημικό-ανοσοενζυματικό μηχανισμό ή φθορισμό. [25]
Mέθοδοι ενίσχυσης ευαισθησίας in situ υβριδισμού
H ενίσχυση της ευαισθησίας του in situ υβριδισμού μπορεί να γίνει σε επίπεδο αναζητούμενης αλληλουχίας προ του υβριδισμού ή σε επίπεδο ανίχνευσης της μετά τον υβριδισμό. H πρώτη στρατηγική επιτυγχάνεται με την αύξηση του αριθμού των αντιγράφων DNA ή RNA με τη βοήθεια ενδοκυττάριας αλυσιδωτής αντίδρασης πολυμεράσης (in situ PCR) και ακολούθως, υβριδισμό των πολλαπλασιασθέντων αλληλουχιών με το δείκτη. H δεύτερη στρατηγική επιτυγχάνεται με μεθόδους ενίσχυσης του σήματος μετά την αντίδραση υβριδισμού, η σπουδαιότερη από τις οποίες είναι αυτή που βασίζεται στην ενζυματική εναπόθεση τυραμιδίου (μέθοδος CARD). H μέθοδος αυτή επιτρέπει τη βελτίωση της ευαισθησίας της αντίδρασης 5-10 φορές, με αποτέλεσμα να καθιστά δυνατή την ανίχνευση ακόμη και ενός αντιγράφου νουκλεοτιδικής αλληλουχίας. [25]
Iδιαίτερα σημαντικές είναι οι εφαρμογές του in situ υβριδισμού στο κλινικό εργαστήριο και αφορούν την ανίχνευση ιών, τις μεταβολές γονιδίων σε επίπεδο πυρηνικού DNA (απώλειες, ενισχύσεις) αλλά και έκφρασης αυτών σε επίπεδο κυτταροπλασματικού mRNA, και τον προσδιορισμό δομικών και αριθμητικών χρωματοσωματικών ανωμαλιών.[25]
Προσδιορισμός γονιδιακών μεταβολών
Aπώλειες ή ενισχύσεις κρίσιμων γονιδίων με καθοριστική σημασία για την πρόγνωση και θεραπευτική αντιμετώπιση νεοπλασμάτων μπορούν να προσδιορισθούν απευθείας εντός του κυττάρου με τεχνική in situ υβριδισμού. H τεχνική μπορεί να εφαρμοσθεί σε μια απλή ιστολογική τομή από μπλοκ παραφίνης νεοπλασματικού ιστού που εστάλη για ιστολογική εξέταση και έχει μονιμοποιηθεί σε ουδέτερη φορμόλη. Ένα από τα κρίσιμα γονίδια με σημαντική προγνωστική και θεραπευτική σημασία στο καρκίνωμα του μαστού είναι αυτό που κωδικοποιεί την πρωτεΐνη c-erbB-2 (ή HER-2 /neu), το οποίο φαίνεται να εμφανίζει ενίσχυση /υπερέκφραση στο 20-30% των περιπτώσεων. H πρωτεΐνη c-erbB-2 εντοπίζεται στην κυτταρική μεμβράνη και είναι μια κινάση τυροσίνης με λειτουργία υποδοχέα έναντι πεπτιδίων που διεγείρουν την κυτταρική αύξηση, διαφοροποίηση και προσκολλητικότητα. Aν και η υπερέκφραση της πρωτεΐνης μέχρι τώρα προσδιορίζεται ανοσοϊστοχημικά, εν τούτοις η τεχνική του in situ υβριδισμού επιτρέπει την άμεση ταυτοποίηση ενίσχυσης του γονιδίου εντός των πυρήνων των καρκινωματωδών κυττάρων και ταυτόχρονα μέτρηση του ποσοστού των νεοπλασματικών κυττάρων που παρουσιάζουν ενίσχυση (Eικόνα 18 ). Aυτό έχει ιδιαίτερη σημασία για τους ασθενείς με προχωρημένο καρκίνωμα μαστού που πρόκειται να επιλεγούν για θεραπεία με Herceptin, ιδίως σε περιπτώσεις αμφιλεγόμενων αποτελεσμάτων του ανοσοϊστοχημικού προσδιορισμού της πρωτεΐνης. Eπίσης, θα πρέπει να ληφθεί υπ' όψιν η υπεροχή της διατήρησης του κυτταρικού DNA σε σχέση με την πρωτεΐνη που μπορεί ευκολότερα να χάσει την αντιγονικότητα της, με αποτέλεσμα ψευδώς αρνητικά αποτελέσματα.[25]

Eικόνα 18 . Eνίσχυση γονιδίου C-ERBB2 σε καρκίνωμα μαστού.[25]
Προσδιορισμός χρωματοσωματικών διαταραχών (FISH)
Oι χρωματοσωματικές ανωμαλίες θεωρήθηκαν από τις αρχές του 20ου αιώνα ως βασικό συστατικό των νεοπλασμάτων και μελετήθηκαν αρχικά με την κλασσική κυτταρογενετική, κυρίως σε αιματολογικά νεοπλάσματα. H κλασσική κυτταρογενετική όμως, λόγω των προβλημάτων επιλογής, δειγματοληψίας και καλλιέργειας των νεοπλασματικών κυττάρων, δεν καθιερώθηκε ως μέθοδος ρουτίνας στους συμπαγείς όγκους. Eναλλακτικά αναπτύχθηκε αργότερα η κυτταρογενετική της μεσόφασης, η οποία βασίζεται στον in situ υβριδισμό αλληλουχιών χρωματοσωμάτων με τη χρήση δεικτών DNA σε ακέραια κύτταρα, χωρίς δηλαδή καλλιέργεια κυττάρων και λήψη μεταφάσεων. Eπειδή οι δείκτες που χρησιμοποιήθηκαν αρχικά ήταν σημασμένοι με φθορίζουσες χρωστικές και η ανίχνευση της αντίδρασης υβριδισμού γινόταν με μικροσκοπία φθορισμού, η τεχνική ονομάσθηκε φθορίζον in situ υβριδισμός (Fluorescent in situ hybridization, FISH). Θα πρέπει να σημειωθεί ότι σήμερα η ίδια τεχνική μπορεί να εφαρμοσθεί με ανίχνευση ενζυματικού τύπου και χρωμογόνα υποστρώματα και ονομάζεται Chromogenic in situ Hybridization (CISH). Tα επιχρίσματα και τα εντυπώματα ακέραιων κυττάρων αποτελούν άριστο υλικό για τη μελέτη των χρωματοσωματικών διαταραχών, αν και ορισμένες από τις εφαρμογές μπορούν αξιόπιστα να γίνουν και σε τομές ιστών μονιμοποιημένες σε φορμόλη και εμπεδωμένες σε παραφίνη. Tο μεγάλο πρόβλημα στις τελευταίες αποτελεί η διαφόρου βαθμού απώλεια χρωματοσωματικού υλικού, λόγω της τυχαίας τομής του πυρήνα κατά τη μικροτόμηση. Σημαντική εφαρμογή της τεχνικής FISH είναι ο προσδιορισμός αριθμητικών διαταραχών που αφορούν είτε σε απώλεια είτε σε αύξηση του αριθμού αντιγράφων ενός χρωμοσώματος (Eικόνα 19). Oρισμένες αριθμητικές ανωμαλίες φαίνεται ότι σχετίζονται σταθερά με συγκεκριμένους τύπους νεοπλασμάτων και αποτελούν βασικό συστατικό της προκαλούμενης ανευπλοειδίας του DNA, η οποία μπορεί να έχει σημαντική προγνωστική αξία 12-14. Aριθμητικές ανωμαλίες στα χρωματοσώματα 13, 18 και 21, συνήθως με τη μορφή της τρισωμίας, χαρακτηρίζουν τα σύνδρομα Patau, Edward και Down αντίστοιχα και αποτελούν σημαντικές εφαρμογές προγεννητικού ελέγχου που μπορούν να απαντηθούν ταχύτατα με τη μέθοδο FISH μεσοφασικών κυττάρων. Tο δεύτερο είδος εφαρμογών της τεχνικής FISH είναι οι δομικές χρωματοσωματικές διαταραχές, οι οποίες μπορούν να ανιχνευθούν με τη χρήση δεικτών έναντι μιας ή περισσότερων ειδικών χρωματοσωματικών αλληλουχιών. Σημαντικές δομικές διαταραχές είναι η διαμετάθεση bcr-abl, γνωστή και ως χρωματόσωμα της Φιλαδέλφειας που αποτελεί διαγνωστικό και προγνωστικό δείκτη της χρόνιας μυελογενούς λευχαιμίας, και η διαμετάθεση t(11;22) (q24;q12), η οποία είναι χαρακτηριστική του σαρκώματος Ewing και των αρχέγονων νευροεξωδερματικών όγκων (PNETs).[25]

Eικόνα 19 . Fish και αριθμητικές χρωματοσωματικές διαταραχές σε καρκίνωμα μαστού[25]
Η έρευνα για την ανίχνευση γονιδιακών μεταλλάξεων στο γονίδιο APC απαιτεί πολύ χρόνο και χρήματα εξαιτίας της ετερογένειας των μεταλλάξεων που ήδη έχουν καταγραφεί. Επιτρέπει, όμως, την αναγνώριση των φορέων παθογόνων μεταλλάξεων, δίνοντας έτσι τη δυνατότητα επισταμένης επίβλεψής τους και της πρώιμης θεραπευτικής επέμβασης. Μέχρι πρόσφατα, οι περισσότερες γενετικές αναλύσεις έχουν γίνει σε ερευνητικό επίπεδο. Τα τελευταία έτη, όμως, παρατηρούμε την εφαρμογή τους σε κλινικό επίπεδο, σε εξειδικευμένα Κέντρα. Οι τεχνικές που χρησιμοποιούνται για την ανίχνευση των μεταλλάξεων, χρησιμοποιούν DNA που μπορεί να προέρχεται από αίμα ή άλλες πηγές. Η ανίχνευση γονιδιακών μεταλλάξεων μπορεί να γίνει με διάφορες τεχνικές. Ανάμεσα στις τεχνικές που χρησιμοποιούνται ευρέως, είναι η μέθοδος της Πολυμορφικής Διαμόρφωσης του Μονόκλωνου DNA (SSCP). Η τεχνική της SSCP εμφανίζεται ως η πλέον κατάλληλη, αφού η ευαισθησία της μεθόδου για τμήματα DNA με μέγεθος 250-350 bp27, όπως αυτά που λαμβάνουμε από τον πολλαπλασιασμό των εξονίων 1-14 του γονιδίου APC, είναι περισσότερο από 80%. Οι αλληλουχίες νουκλεοτιδίων των προϊόντων της PCR, που εμφανίζουν μη φυσιολογική ηλεκτροφορητική κινητικότητα στην SSCP, προσδιορίζονται με τη βοήθεια της τεχνικής "Sequencing". Η ίδια τεχνική μπορεί να εφαρμοστεί σε DNA που έχει προέλθει από διάφορες πηγές (νωπό ή κατεψυγμένο αίμα ή ιστό εγκλεισμένο σε μπλοκ παραφίνης). Η τεχνική της ανίχνευσης Ελλιπών Μορίων Πρωτεΐνης (ΡΙΤ), όπου γίνονται in vitro μεταγραφή και μετάφραση του γονιδιακού υλικού και έχει εφαρμοστεί σε ασθενείς με APC 28, κυρίως για το εξόνιο 15, η οποία έχει το μειονέκτημα ότι ανιχνεύει μόνο τις μεταλλάξεις εκείνες που καταλήγουν σε μια ελλιπούς μεγέθους πρωτεΐνη.[26]
Η NIRCA ανάλυση υιοθετήθηκε για να προσδιορίσει διαφορές στις ακολουθίες μεταξύ RT-PCR προϊόντων και κλώνου πλασμιδίων pRT14. Το RT-PCR προϊόν παρήχθη από FVB και C57BL/6 RNA από συκώτι όπως περιγράφεται με χρήση ολιγο-dT για να εκκινήσει την σύνθεση του πρώτου κλώνου. Τα RT-PCR προϊόντα, το πλασμίδιο pRT14 DNA και γενωμικό DNA από FVB ποντίκια ενισχύθηκαν με χρήση εκκινητών που περιείχαν T7 (κωδικές) ή SP6 (αντικωδικές) εκκινητικές αλληλουχίες. Τα τμήματα του εκκινητή (όλα 5´à3´) τρέχουν από την 63 βάση στην 901
(TF63, TAATACGACTCACTATAGGATGGCTGCTGCCT
SR901, ATTTAGGTGACACTATAGAAGGCTCCCACACG) ή από τη βάση 705 στην 1324 (TF705, TAATACGACTCACTATAGGAGATAGCGTCCG
SR1324, ATTTAGGTGACACTATAGAATTACTTGGTGACCTG).
Κάθε ένα από τα προϊόντα της pcr αντίδρασης χρησιμοποιήθηκε ως πρότυπο για RNA παραγωγή με χρήση T7 και SP6 πολυμεράσες στις χωριστές αντιδράσεις. Όλοι οι συνδυασμοί κωδικών και αντικωδικών προϊόντων υβριδοποιήθηκαν, και έγινε πέψη τους με δύο διαφορετικά μίγματα RNAασών. Ο πρώτος ήταν ένα μίγμα από 1/300 RNAση ![]() 1 και 1/100 RNAse
1 και 1/100 RNAse ![]() 2, και ο δεύτερος ήταν 1/100 RNAση
2, και ο δεύτερος ήταν 1/100 RNAση ![]() 3. Μετά από την πέψη, οι αντιδράσεις διαχωρίστηκαν σε ένα 2% πήκτωμα αγαρόζης και φωτογραφήθηκαν.(Εικόνα 20) [27]
3. Μετά από την πέψη, οι αντιδράσεις διαχωρίστηκαν σε ένα 2% πήκτωμα αγαρόζης και φωτογραφήθηκαν.(Εικόνα 20) [27]

Εικόνα 20 NIRCA αποτελέσματα [28]
Tεχνολογία microchip
H σπουδαιότερη επιστημονική επανάσταση των πρόσφατων χρόνων είναι η ανάπτυξη μικροτσίπ εφοδιασμένων με ολιγονουκλεοτίδια που επιτρέπουν ταχύτατες και ευρείας κλίμακας μελέτες γονιδίων. Tα συστήματα αυτά με βάση την αντίδραση υβριδισμού επιτρέπουν την ανάλυση αλληλουχίας μεγάλου αριθμού επιλεγμένων γονιδίων και τον προσδιορισμό της έκφρασης αυτών σε επίπεδο mRNA. H έκφραση επομένως χιλιάδων γονιδίων ταυτόχρονα μπορεί να μελετηθεί ποσοτικά και με ακρίβεια με τη βοήθεια ηλεκτρονικού υπολογιστή, εξασφαλίζοντας έτσι υψηλή ποιότητα και επαναλληψιμότητα.[28]
[1] http://www.au-kbc.org/research_areas/bio/projects/bcinfo/index.html
[2] http://www.ich.ucl.ac.uk/cmgs/oligohyb.htm
[3] http://las.perkinelmer.com/content/snps/protocol.asp
[4] http://www.ich.ucl.ac.uk/cmgs/ptt98.htm
[5] BB310: Molecular Genetics , Lecture 22: Applications & Examples: Genetic Testing, Dr Eve Lutz ,Department of Bioscience
[6] Combined SSCP/duplex analysis by capillary electrophoresis for more efficient mutation detection., Kozlowski P, Krzyzosiak WJ., Laboratory of Cancer Genetics, Institute of Bioorganic Chemistry, Polish Academy of Sciences, Noskowskiego 12/14, 61-704 Poznan, Poland., PMID: 11452040
[7] http://en.wikipedia.org/wiki/Single_strand_conformation_polymorphism
[8] Created: 1st December 1999, last updated: 21st February 2000, © 1999 ABRF, METHODS AND REVIEWS, Considerations in Adding Mutation Detection Services to a Sequencing Core Facility, M. Roxane Bonnera and Linda Wood Ballardb aLaboratory of Molecular Systematics and Evolution, University of Arizona, Tucson, AZ; bGenomics Core Facility, Huntsman Cancer Institute, University of Utah, Salt Lake City, UT
[9] http://www.amonline.net.au/evolutionary_biology/images/sscp.gif
[10] Combined SSCP/duplex analysis by capillary electrophoresis for more efficient mutation detection. Kozlowski P, Krzyzosiak WJ. Laboratory of Cancer Genetics, Institute of Bioorganic Chemistry, Polish Academy of Sciences, Noskowskiego 12/14, 61-704 Poznan, Poland., PMID: 11452040
[11] http://www.uark.edu/campus-resources/mivey/m4233/heterod.gif
[12] Applications of microarray technology in breast cancer research. Cooper CS,The Institute of Cancer Research, Sutton,
[13]http://promesip.med.auth.gr/pathology/modules.php?op=modload&name=UpDownload&file=index&req=getit&lid=86
[14] http://www.fao.org/DOCREP/003/X6884E/x6884e00.jpg
[15] http://www.biocompare.com/images/techart/Bulletin_2408_2.jpg
[16] http://en.wikipedia.org/wiki/Temperature_gradient_gel_electrophoresis
[17] http://documents.plant.wur.nl/pri/biointeractions/images/slide14.jpg
[18] http://www.microbac.com/technical_articles/news_detail.php?news_ID=7
[19] http://bipar.vet-alfort.fr/thematiques/890/gal/principe_tgge.jpg
[20] Cell. 1979 Jan, Vol 16, pages 191-200; Length-independent separation of DNA restriction fragments in two-dimensional gel electrophoresis.
[21] M. Roxane Bonnera and Linda Wood Ballardb, METHODS AND REVIEWS, Considerations in Adding Mutation Detection Services to a Sequencing Core Facility, , aLaboratory of Molecular Systematics and Evolution, University of Arizona, Tucson, AZ; bGenomics Core Facility, Huntsman Cancer Institute, University of Utah, Salt Lake City, UT, Created: 1st December 1999, last updated: 21st February 2000, 1999 ABRF
[22] Jarmo Körkkö, Susanna Annunen, Tero Pihlajamaa, Darwin J. Prockop, and Leena Ala-Kokko, Genetics, Conformation sensitive gel electrophoresis for simple and accurate detection of mutations: Comparison with denaturing gradient gel electrophoresis and nucleotide sequencing, Center for Gene Therapy, Allegheny University of the Health Sciences, MCP-Hahnemann School of Medicine, Philadelphia, PA 19102; and Collagen Research Unit, Biocenter and Department of Medical Biochemistry, University of Oulu, SF-90220 Oulu, Finland Vol. 95, Issue 4, 1681-1685, February 17, 1998
[23] Graham R. Bignell, William Warren, Sheila Seal, Meiko Takahashi, Elizabeth Rapley, Rita Barfoot, Helen Green, Carolanne Brown, Patrick J. Biggs, Sunil R. Lakhani, Christopher Jones, Juliana Hansen, Edward Blair, Benedikt Hofmann, Reiner Siebert, Gwen Turner, D. Gareth Evans, Connie Schrander-Stumpel, Frits A. Beemer, Ans van den Ouweland, Dicky Halley, Bertrand Delpech, Mark G. Cleveland, Irene Leigh, Jaakko Leisti, Sonja Rasmussen, Margaret R. Wallace, Christiane Fenske, Piu Banerjee, Naoki Oiso, Ranbir Chaggar, Samantha Merrett, Niamh Leonard, Marcel Huber, Daniel Hohl, Pam Chapman, John Burn, Sally Swift, Anna Smith, Alan Ashworth & Michael R. Stratton, Identification of the familial cylindromatosis tumour-suppressor gene, Nature Genetics 25, 160 - 165 (2000) doi:10.1038/76006
[24] Gloria A. Thomas1, Diana L. Williams2 and Steven A. Soper1a, Capillary Electrophoresis-based Heteroduplex Analysis with a Universal Heteroduplex Generator for Detection of Point Mutations Associated with Rifampin Resistance in Tuberculosis, 1 Chemistry Department, Louisiana State University, Baton Rouge, LA 70803. 2 Molecular Biology Research Department, Laboratory Research Branch, National Hansen’s Disease Programs at the School of Veterinary Medicine, Louisiana State University, Baton Rouge, LA 70803, Clinical Chemistry 47: 1195-1203, 2001.
[25] KAΠPANOΣ NIKHΦOPOΣ, In situ υβριδισμός. Mια μοναδική μορφολογική μέθοδος μοριακής βιολογίας με πολύτιμες κλινικές εφαρμογές, Iατρός Παθολογοανατόμος, Mονάδα Mοριακής Παθολογικής Aνατομικής, NΓNM Aμαλία Φλέμιγκ.
[26] http://www.digestiveoncology.org.gr/volume2_3_3.asp
[27] Michael W. BRADBURY1 and Paul D. BERK, Mitochondrial aspartate aminotransferase: direction of a single protein with two distinct functions to two subcellular sites does not require alternative splicing of the mRNA, Department of Medicine (Division of Liver Diseases), Mount Sinai School of Medicine, Box 1633, 1 Gustave L. Levy Place, New York, NY 10029, U.S.A. Biochem. J. (2000) 345 (423–427).
[28] Basset DE Jr, Eisen MB, Boguski MS. Gene expression informatics½it£s all in your mine. Nat Genet 1999; 21:51-55
14. Ανίχνευση μοριακών δεικτών καρκίνου και καρδιακών νοσημάτων με μεθόδους Μοριακής Διαγνωστικής.
Κατά τη διάρκεια των προηγούμενων 20 ετών, οι τεχνολογικές πρόοδοι στη μοριακή βιολογία κατέστησαν ανεκτίμητες στην κατανόηση της παθογένεσης του ανθρώπινου καρκίνου. Η εφαρμογή της μοριακής τεχνολογίας στη μελέτη του καρκίνου δεν έχει μόνο προσφέρει στην πρόοδο της διάγνωσης των όγκων, αλλά έχει παράσχει επίσης δείκτες για το την αξιολόγηση της πρόγνωσης και την εξέλιξη των ασθενειών. Ο στόχος της μοριακής ανάλυσης του καρκίνου είναι να παράσχει μια περιεκτική συλλογή από πολύ περισσότερες up-to-date τεχνικές για την ανίχνευση των μοριακών αλλαγών στον ανθρώπινο καρκίνο. Κορυφαίοι ερευνητές στον τομέα έχουν συμβάλει με πρακτικές διαδικασίες σε ένα ευρύ φάσμα από τεχνικές. Η Μοριακή ανάλυση του καρκίνου περιλαμβάνει κεφάλαια που περιγράφουν τεχνικές για τον προσδιορισμό χρωμοσωμικών ανωμαλιών και περιλαμβάνουν: επιτόπια υβριδοποίηση φθορισμού (FISH), φασματική καρυοτύπηση (SKY), συγκριτική γενωμική υβριδοποίηση (CGH), και ανάλυση μικροδορυφόρων. Η FISH έχει έναν προεξέχοντα ρόλο στη μοριακή ανάλυση του καρκίνου και μπορεί να χρησιμοποιηθεί για την ανίχνευση αριθμητικών και δομικών χρωμοσωμικών ανωμαλιών. Πρόσφατα περιγραφής SKY, στο οποίο όλα τα ανθρώπινα χρωμοσώματα μετάφασης απεικονίζονται με συγκεκριμένα χρώματα, επιτρέπει τον καθορισμό όλων των χρωμοσωμικών αναδιοργανώσεων και δεικτών χρωμοσωμάτων σε ένα καρκινικό κύτταρο. Πρωτόκολλα για την ανίχνευση χρωμοσωμικών αναδιοργανώσεων με PCR και RT-PCR περιγράφονται, καθώς επίσης και η τεχνική DNA δακτυλοσκοπία, ένα ισχυρό εργαλείο για σωματικές γενετικές αλλαγές στην καρκινογένεση. Διάφορες προσεγγίσεις για να προσδιορίσουν τις μεταλλαγές περιγράφονται λεπτομερώς, και περιλαμβάνουν SSCP, DGGE, μη ισοτοπική RNase δοκιμή διάσπασης, η πρωτεϊνική δοκιμή αποκοπής, και DNA αλληλούχιση. Μια αλλαγή στη θέση μεθυλίωσης του DNA παρατηρείται συνήθως στον καρκίνο, και συγκεκριμένη μεθοδολογία για την ανάλυση μεθυλίωσης αναλύεται παρακάτω. Η ανάλυση της έκφρασης γονιδίων αντιπροσωπεύει μια βασική περιοχή από την έρευνα στη μελέτη του ανθρώπινου καρκίνου. Σφαιρική ανάλυση RNA έκφρασης χρησιμοποιώντας τεχνολογία μικροσυστιχιών επιτρέπει τον προσδιορισμό των γονιδίων που είναι διαφορικά εκφρασμένα στον όγκο έναντι των κανονικών ιστών. Αυτό είναι μια ισχυρή προσέγγιση για προσδιορίσουμε τα γονίδια που παίζουν κεντρικό ρόλο στην ανάπτυξη της ασθένειας ή στην πρόοδό της και μπορούμε επίσης να προσδιορίσουμε νέους διαγνωστικούς δείκτες.[1]
Μια μείωση του μήκους των τελομερών, μαζί με την έκφραση από το ένζυμο συντήρησης των τελομερών, την τελομεράση, έχει περιγραφεί σε ένα ευρύ φάσμα από τους ανθρώπινους καρκίνους. Η περιγραφή της μέτρησης του μήκους του τελομερούς και των επιπέδων της τελομεράσης, είναι ένας τομέας εκτενούς μελέτης στον τομέα της έρευνας του καρκίνου. [1]
Ο καρκίνος είναι μια σύνθετη ασθένεια που εμφανίζεται ως αποτέλεσμα μιάς προοδευτικής συσσώρευσης από τις γενετικές παρεκκλίσεις και επιγενετικές αλλαγές που επιτρέπει τη διαφυγή από φυσιολογικό κυτταρικό και περιβαλλοντικό έλεγχο. Τα νεοπλαστικά κύτταρα μπορούν να έχουν πολυάριθμες επίκτητες γενετικές ανωμαλίες συμπεριλαμβανομένης της ανευπλοειδίας, χρωμοσωμικές αναδιοργανώσεις, ενισχύσεις, διαγραφές, αναδιοργανώσεις γονιδίων, και απώλεια λειτουργίας ή ενίσχυση λειτουργίας μεταλλαγές. Οι πρόσφατες μελέτες τονίζουν επίσης τη σημασία των επιγενετικών αλλαγών σε ορισμένα γονίδια με αποτέλεσμα την αδρανοποίηση από τις λειτουργίες τους σε μερικούς ανθρώπινους καρκίνους. Αυτή η παρέκκλιση δείχνει κοινή ανώμαλη συμπεριφορά σε όλα τα νεοπλαστικά κύτταρα: δυσρυθμιζόμενη αύξηση, έλλειψη παρεμπόδισης επαφών, γενωμική αστάθεια, και ροπή για μετάσταση. Τα γονίδια που επηρεάζονται από τις μεταλλαγές στον καρκίνο μπορεί να διαιρεθούν σε δύο κύριες κατηγορίες: μεταλλαγές σε γονίδια που έχουν το κέρδος στη λειτουργία (ενεργοποίηση), που είναι γνωστά ως ογκογονίδια, και μεταλλαγές σε γονίδια για τα οποία και τα δύο αλληλόμορφα έχουν χάσει την λειτουργία τους (αδρανοποίηση), που είναι γνωστά ως γονίδια καταστολής όγκων. Κοντά στα 100 γονίδια έχουν παρουσιαστεί για να διαδραματίσουν ένα ρόλος στην ανάπτυξη ή την πρόοδο στους ανθρώπινους καρκίνους, μερικοί των οποίων έχουν εμπλακεί σε ένα ευρύ φάσμα από κακοήθης όγκους, εκτιμώντας ότι άλλοι είναι μοναδικοί σε έναν συγκεκριμένο τύπο. Οι καρκίνοι μπορούν να προκύψουν μέσω μιάς παρέκκλισης από διαφορετικούς συνδυασμούς γονιδίων, που μπορούν στη συνέχεια να αλλοιώνονται, να υπερεκφράζονται, ή να διαγράφονται. Η τάξη στην οποία αυτά τα γεγονότα εμφανίζονται έχει αποδειχθεί επίσης ότι είναι σημαντική. Παραδείγματος χάριν, στον καρκίνο του μαστού έχει προταθεί ότι τουλάχιστον 10 ευδιάκριτες αλλαγές γονίδιων μπορούν να περιληφθούν στην έναρξη και πρόοδο της ασθένειας. Η μελέτη του καρκίνου του κόλου έχει παρουσιάσει την καρκινιγένεση σαν μια πολυβάθμια διαδικασία με ανάμειξη της ενεργοποίησης των κυτταρικών ογκογονιδίων, τη διαγραφή των πολλαπλών χρωμοσωμικών περιοχών, και την απώλεια λειτουργίας από τα γονίδια καταστολής όγκων. Πρόοδοι της τεχνολογίας στη μοριακή βιολογία τα τελευταία 20–25 χρόνια έχουν οδηγήσει σε μια δραματική αύξηση στον προσδιορισμό των μοριακών διαδικασιών της βολβοειδούς καρκινογένεσης. Κατά τη διάρκεια αυτής της περιόδου, η μοριακή βάση του καρκίνου no εμπεριέχει το μυστήριο που είχε κάποτε. Είναι, εντούτοις, επίσης σαφές ότι η τεχνογνωσία που έχει συσσωρευθεί είναι ανεπαρκής να απαιτήσει μια συνολική κατανόηση του μηχανισμού της ανάπτυξης του καρκίνου.[1]
Η μεταλλακτική ανάλυση μπορεί να εκτελεσθεί με χρησιμοποίηση ποικίλων τεχνικών, και η πλειοψηφία αυτών τονίζεται παρακάτω. Η ενίσχυση των συγκεκριμένων περιοχών DNA η RNA με PCR έχει ανοίξει ατελείωτες δυνατότητες που μπορούν να χρησιμοποιηθούν για γρήγορη και αποδοτική ανίχνευση των αλλαγών, ακόμη και απλών αλλαγών νουκλεοτιδίων. Αυτή η PCR- τεχνική είναι βασισμένη στις αλλαγές στην ηλεκτροφορητική κινητικότητα προκληθείσα από αλλαγμένη δευτεροταγή δομή της απλής αλυσίδας (single-strand conformation polymorphism), από τα αλλαγμένα ποσοστά διαχωρισμού τμημάτων του DNA (denaturing gradient gel electrophoresis), η με RNase cleavage assays.Η PCR μπορεί ακόμη να χρησιμοποιηθεί για γρήγορή και ποσοτική ανίχνευση από χρωμοσωμικές αναδιοργανώσεις, όπως συνήθως παρατηρείται στη λευκαιμία.Η PCR σχεδιάστηκε για να ενισχύσει συγκεκριμένα γενωμικά τμήματα που δεν είναι κανονικά εφαπτόμενα και είναι, επομένως, μοναδικά σε αυτόν τον τύπο αναδιοργάνωσης γονιδίων. Μετατροπή του RNA σε DNA με την αντίστροφη μεταγραφάση (RT) πριν από την PCR απαιτείται συνήθως για την ανάλυση. Εντούτοις, σε μερικές περιπτώσεις, γενωμικού DNA μπορούμε να χρησιμοποιήσουμε για την άμεση ενίσχυση σημείων σπασιμάτων από μετατόπιση. Μια παραλλαγή PCR εμπεριέχει τη χρήση DNA δακτυλικών αποτυπωμάτων για να ανιχνεύσει γενετικά αίτια στον καρκίνο. Οι εκκινητές είναι συχνά αυθαίρετοι ή επαναλαμβάνουν (π.χ., ALU) ακολουθίες, που θα δώσουν, μετά από την ηλεκτροφόρηση, ένα δακτυλικό αποτύπωμα DNA που μπορεί να χρησιμοποιηθεί για την ανίχνευση γενετικών ανωμαλιών. Επαναλήψεις μικροδορυφόρων εμφανίζονται σε όλο το γονιδίωμα και μπορεί να χρησιμοποιηθούν ως δείκτες για γενετικές αλλαγές, συνήθως για την απώλεια ετεροζυγωτίας, που θα δείξει ότι μια διαγραφή έχει εμφανιστεί να επικαλύπτει τον συγκεκριμένο δείκτη. Για τα συγκεκριμένα γονίδια σχετικά με ορισμένους καρκίνους, η μεταλλακτική ανάλυση μπορεί να πραγματοποιηθεί με χρησιμοποίηση ενός δείκτη στη ανάλυση αποκοπής. Αυτή η ανάλυση περιλαμβάνει τον προσδιορισμό ανώμαλών πολυπεπτιδίων που συντίθενται in vitro από RT-PCR προϊόντα, και οι μεταλλαγές περικοπής συνήθως επιβεβαιώνονται με ανάλυση ακολουθίας [1]
Ανάλυση έκφρασης RNA
Η τεχνολογία μικροσυστοιχιών DNA, που χρησιμοποιεί την ανάλυση δείκτη υψηλής πυκνότητας δύο διαστάσεων ολογονουκλεοτιδίων περιέχει εκατοντάδες ή χιλιάδες δείκτες, ολογονουκλεοτιδίων αντιπροσωπεύει ένα νέα ισχυρό εργαλείο στην ανάλυση ακολουθίας DNA για ποικίλες γενετικές μεταλλαγές Υβριδοποίηση toc DNA μικροσυστοιχιών επιτρέπει την ταυτόχρονη παράλληλη ανάλυση έκφρασης από χιλιάδες γονίδια. Για γονίδιο με έκφραση υψηλής-ρυθμοαπόδοσης ο σχεδιάσμός περιγράμματος όλο και περισσότερο αποτελεί μια πολύτιμη μέθοδο για να προσδιοριστεί γονίδια διαφορικά εκφρασμένα σε όγκο vs κανονικών ιστών. Έκφραση γονιδίων μικροσυστοιχιών υπόσχεται πολλά για τις μελέτες της ανθρώπινης καρκινογένεσης, και τα πολλά στοιχεία έκφρασης των γονιδίων τα συνολικά παραχθέντα έχουν τη δυνατότητα να παρέχουν νέες ιδέες στη θεμελιώδη βιολογία του καρκίνου σε μοριακό επίπεδο. Πράγματι, η τεχνολογία μικροσυστοιχιών του cDNA έχει αρχίσει ήδη να βοηθά στη διευκρίνιση των γενετικών γεγονότων που κρύβονται κάτω από την έναρξη και την εξέλιξη μερικών ανθρώπινων καρκίνων. Τα διαφορικά εκφρασμένα γονίδια μπορούν επίσης να ανιχνευθούν από άλλες τεχνικές όπως η διαφορική επίδειξη, που περιλαμβάνει μία τυχαίου εκκινητή RT-PCR επίδειξη δακτυλοσκόπησης από τα υποσύνολα εκφρασμένου RNA, ή αφαιρετική υβριδοποίηση, που περιλαμβάνει τον εμπλουτισμό των γονιδίων κατά προτίμηση των εκφρασμένων σε έναν ιστό έναντι ενός δεύτερου.[1]
Fluorescence in situ hybridization (FISH) είναι μια από τις τεχνικές με σημαντικό ρόλο στην μοριακή ανάλυση του καρκίνου. Μπορεί να χρησιμοποιηθεί για την απλή ανίχνευση αριθμητικών και δομικών χρωμοσωμικών ανωμαλιών αυτός μπορεί να εμφανιστεί στα καρκινικά κύτταρα και είναι ιδιαίτερα χρήσιμη ως εργαλείο για τη διάγνωση μη τυχαίων μετατοπίσεων στη λευχαιμία και σε πολυάριθμους άλλους καρκίνους. Μέχρι σήμερα, οι περισσότερες FISH μελέτες έχουν περιλάβει τη χρήση ολόκληρου του χρωμοσώματος γονιδιακών δεικτών. Αυτό έχει γίνει σε νέα επίπεδα με την ανάπτυξη φασματικής καρυοτύπησης, που περιλαμβάνει την υβριδοποίηση 24 ιχνηθετημένων με φθορίζουσες ουσίες χρωμοσωμικών χρωματισμένων δεικτών που στη μετάφαση δινει ταυτόχρονη απεικόνιση σε κάθε ένα από τα χρωμοσώματα με ένα διαφορετικό χρώμα. Χρησιμοποιώντας αυτήν την μέθοδο, είναι δυνατό να καθορίσουμε όλες τις χρωμοσωμικές αναδιοργανώσεις και να προσδιορίσουμε τον χρωμοσωμικό δείκτη στα καρκινικά κύτταρα. Συγκριτική γενομική υβριδοποίηση (CGH) είναι μια FISH-βασιζόμενη τεχνική που μπορεί να ανιχνεύσει τα κέρδη και τις απώλειες ολόκληρων των χρωμοσωμάτων και υποχρωμοσωμικές περιοχές. Η CGH είναι βασισμένη σε διχρωματική, ανταγωνιστική FISH με διαφορικά ιχνηθετημένο όγκος και DNA αναφοράς στα κανονικά χρωμοσώματα της μετάφασης και μπορεί να ανιχνεύσει ολόκληρο το γονιδίωμα χωρίς προγενέστερη γνώση από τις συγκεκριμένες χρωμοσωμικές ανωμαλίες.[1]
Μερικές μοριακές μέθοδοι θα αναλύσουν συγκεκριμένες αλλαγές στη δομή του DNA η σε γενωμικές τροποποιήσεις. Αλλαγές στη θέση μεθυλίωσης του DNA είναι μια από τις πιό κοινές ανιχνεύσιμες ανωμαλίες στον ανθρώπινο καρκίνο. Υπερμεθυλίωση μέσα σε υποκινητές από επιλεγμένα γονίδια είναι ιδιαίτερα κοινή και είναι συχνα συνδεδεμένη με αδρανοποίηση του περιληφθέντος γονιδίου ή γονιδίων και μπορεί να είναι ένα πρόωρο γεγονός στην παθογένεση μερικών καρκίνων, εκτιμώντας ότι άλλα γονίδια μεθυλιώνονται κατά τη διάρκεια της εξέλιξης των ασθενειών [1].
Τα τελομερή είναι επαναλαμβανόμενες ακολουθίες DNA στις άκρες των χρωμοσωμάτων, που είναι απαραίτητοι για τη διατήρηση της χρωμοσωμικής ακεραιότητας. Μια μείωση του μήκους των τελομερών έχει περιγραφεί σε ένα ευρύ φάσμα των ανθρώπινων καρκίνων, και συμπεριλαμβανομένων των στερεών όγκων και των λευχεμιών. Το ένζυμο της τελομεράσης συνθέτει de novo τελομερικές επαναλήψεις και τις ενσωματώνει επάνω στα 3' άκρα του DNA από τα χρωμοσώματα.Το κόντεμα στα τελομερή στα κανονικά κύτταρα είναι ένα αποτέλεσμα της αντιγραφής του DNA, και μείωση πέρα από ένα κρίσιμο μήκος είναι ένα σήμα για κυτταρική γήρανση. Εντούτοις, η συντήρηση του μήκους των τελομερών, από την ενεργοποίηση του ενζύμου της τελομεράσης, είναι πιθανά ουσιαστική για την αποφυγή του ανθρώπινου καρκίνου των κυττάρων για να αντισταθμίσουν την απώλεια του DNA από τις άκρες των χρωμοσωμάτων. Επομένως, το μέτρημα του μήκους των τελομερών και των επιπέδων της δραστηριότητας του ενζύμου της τελομεράσης είναι σημαντικό στον έλεγχο της προόδου των ασθενειών και στην ανταποκριση της θεραπείας. Πρόσφατα, ο πιθανός χειρισμός της τελομεράσης έχει παραγάγει κάποιο ενθουσιασμό σαν αντικαρκινική στρατηγική.[1]
Κλωνική προέλευση του καρκίνου (Clonal Origin of Cancer)
Οι μέθοδοι που περιγράφηκαν επιτρέπουν στον ερευνητή για να μελετήσει τη μυριάδα από τις γενετικές αλλαγές που μπορούν να εμφανιστούν κατά τη διάρκεια της έναρξης, ανάπτυξης, και εξέλιξης του καρκίνου. Εντούτοις, είναι επίσης δυνατό να παρέχει διορατικότητα στη μετάβαση από τη μεταλλαγή σωματικών κυττάρων στη νεοπλασία. Η κλωνική προέλευση της κυτταρικής ανίχνευσης αξιολογείται στους ασθενείς με πολυμορφισμούς συνδεμένους με το X χρωμόσωμα για να εκμεταλλευθεί την τυχαία αδρανοποίηση του X χρωμοσώματος. Η αδρανοποίηση συσχετίζεται με διαφορικά μεθυλιωμένα σχήματα στα ενεργά και ανενεργά χρωμοσώματα Χ.[1]
Καρκινικοί δείκτες ή δείκτες όγκου είναι φυσιολογικά κυτταρικά συστατικά τα οποία απελευθερώνονται σε παθολογικές ποσότητες από τα ίδια τα νεοπλασματικά κύτταρα από τα κύτταρα του ξενιστή οργανισμού σε απάντηση ενός αντιγονικού (νεοπλασματικού) ερεθίσματος. Είναι ένζυμα, ορμόνες, ογκοεμβρυικά αντιγόνα και άλλες πρωτείνες Ανιχνεύονται με τη βοήθεια ειδικών τεχνικών στο αίμα, πλάσμα και άλλα υγρά του σώματος (ΕΝΥ, πλευριτικό υγρό, περιτοναικό υγρό, ούρα) σε κύτταρα και ιστούς.
Κανένας δείκτης δεν είναι απόλυτα ειδικός και η ευαισθησία ποικίλλει
Ανιχνεύονται σε χαμηλά επίπεδα σε φυσιολογικές καταστάσεις μη νεοπλασματικές παθήσεις. [2]
διάγνωση
Η μεγάλη συμβολή της ανοσοϊστοχημείας στην παθολογοανατομική διάγνωση αποδίδεται σε τρεις παράγοντες
1. τη διαθεσιμότητα πολλών αντισωμάτων κατάλληλων για χρήση σε αρχειακό υλικό
2. την πρόοδο των τεχνικών ανάκλησης των αντιγόνων (antigen retrieval), η οποία οδηγεί σε αξιόπιστα αποτελέσματα και
3. την ανάπτυξη ευαίσθητων συστημάτων ανίχνευσης
Το μέλλον της ανοσοϊστοχημείας διαγράφεται ελπιδοφόρο, καθώς αναπτύσσονται νέες τεχνικές όπως οι μικροσυστοιχίες ιστού (tissue microarrays) (Εικόνα 1)οι οποίες επιτρέπουν την ανίχνευση μιας συγκεκριμένης πρωτεΐνης σε μεγάλο αριθμό δειγμάτων ή την ανίχνευση μεγάλου αριθμού πρωτεϊνών σε ένα μόνο δείγμα, με σχετικά χαμηλό κόστος.[2]
![]()
Εικόνα 1.[2]
· Στη διερεύνηση των καρκινωμάτων αγνώστου πρωτοπαθούς εστίας ιδιαίτερα χρήσιμη είναι η ταυτοποίηση των εκφραζόμενων κυτοκερατινών.
· Έχουν αναγνωριστεί 20 υπότυποι της κυτοκερατίνης, που έχουν ποικίλα μοριακά βάρη και εκφράζονται διαφορετικά σε κάθε τύπο επιθηλιακών κυττάρων και στα νεοπλάσματα.
· Μονοκλωνικά αντισώματα εναντίον ειδικών υποτύπων των κυτοκερατινών έχουν χρησιμοποιηθεί για να ταξινομηθούν νεοπλάσματα ανάλογα με τη θέση προέλευσής τους
Οι πιο χρήσιμες χρώσεις είναι για τις CK7 (Εικόνα 2)και CK20 (Εικόνα 3)
Η CK20 είναι χαμηλού μοριακού βάρους και εκφράζεται φυσιολογικά στο επιθήλιο του γαστρεντερικού, στο ουροθήλιο και στα κύτταρα Merkel.
Η CK7 εκφράζεται στα νεοπλάσματα του πνεύμονα, της ωοθήκης, του ενδομητρίου και του μαστού.
Αν και κανένας ανοσοφαινότυπος δεν είναι παθογνωμονικός ο ανοσοφαινότυπος CK7-/CK20+ εισηγείται πρωτοπαθή όγκο του παχέος εντέρου ενώ ο ανοσοφαινότυπος CK7+/CK20- περιορίζει τη διαφορική διάγνωση σε καρκινώματα του πνεύμονα, μαστού, χοληφόρων, παγκρέατος, ωοθηκών και ενδομητρίου
![]()
Εικόνα 2 CK7 σε αδενοκαρκίνωμα πνεύμονα [2]
![]()
Εικόνα 3 CK20 σε αδενοκαρκίνωμα παχέος εντέρου [2]
Χρήσιμοι δείκτες για τη διάγνωση νεοπλασμάτων του θυρεοειδούς αδένα είναι οι CK19 (Εικόνα 4 ) και η γκαλεκτίνη (galectin-3).H CK19 εκφράζεται έντονα στα νεοπλασματικά κύτταρα του θηλώδους καρκινώματος ΔΔ:
· καλοήθη αλλοίωση με θηλώδη μορφολογία
· θυλακιώδες καρκίνωμα (η θυλακιώδης ποικιλία του
· θηλώδους καρκινώματος)
![]()
Εικόνα 4 CK19 σε θηλώδες καρκίνωμα θυρεοειδούς αδένα [2]
H γκαλεκτίνη (galectin-3) (Εικόνα 5 )εκφράζεται στα νεοπλασματικά κύτταρα του θυλακιώδους καρκινώματος ΔΔ:
· θυλακιώδες αδένωμα από θυλακιώδες καρκίνωμα (minimally invasive follicular carcinoma) ελλείψει σαφών ιστολογικών κριτηρίων
· θυλακιώδες αδένωμα από θυλακιώδη ποικιλία του θηλώδους καρκινώματος
![]()
Εικόνα 5 galectin-3 σε θυλακιώδες καρκίνωμα θυρεροειδούς [2]
Άλλοι χρήσιμοι δείκτες είναι:
· ο TTF-1 (thyroid transcription factor) (Εικόνα 6 )που εκφράζεται σε καρκινώματα πνεύμονα και θυρεοειδούς
![]()
Εικόνα 6 TTF-1 σε καρκίνωμα πνεύμονα [2]
· η GCDFP (gross cystic disease fibrous protein), η οποία είναι δείκτης αποκρινούς διαφοροποίησης και εκφράζεται σε καρκινώματα μαστού και
· η ουροπλακίνη ΙΙΙ (UROIII), η οποία εισηγείται ουροθηλιακή προέλευση του νεοπλάσματος.
Διάγνωση συχνών νεοπλασμάτων επιθηλιακής προέλευσης, όπως το καρκίνωμα του μαστού Ο έλεγχος της έκφρασης της E-cadherin (Εικόνα 7 )για τη διαφορική διάγνωση μεταξύ πορογενούς και λοβιακού καρκινώματος.
![]()
Εικόνα 7 E-καντχερίνη σε in situ αδενοκαρκίνωμα μαστού [2]
Η εφαρμογή ανοσοχρώσεων κυτοκερατίνης για την ανίχνευση μεταστάσεων στο λεμφαδένα φρουρό
Διάγνωση των νεοπλασμάτων μεσεγχυματικής αρχής, τα οποία χαρακτηρίζονται από μεγάλη ετερογένεια, με ποικίλες κατευθύνσεις διαφοροποίησης και συχνές δυσκολίες στη διαφορική διάγνωση από καλοήθεις ψευδονεοπλασματικές αλλοιώσεις κακοήθη νεοπλάσματα μη μεσεγχυματικής αρχής.[2]
Η αναζήτηση των καρκινικών δεικτών θεωρείται απαραίτητη για τη διάγνωση, γιατί ο ανοσοφαινότυπος αποτελεί μέρος της ταυτότητας του όγκου. Τέτοια νεοπλάσματα είναι:
· το ραβδομυοσάρκωμα
· το επιθηλιοειδές σάρκωμα
· το διαυγοκυτταρικό σάρκωμα
· ο δεσμοπλαστικός στρογγυλοκυτταρικός όγκος και
· οι στρωματικοί όγκοι του γαστρεντερικού
Οι πιο χρήσιμοι δείκτες για την αρχική προσέγγιση του νεοπλάσματος είναι:
Επιθηλιακοί δείκτες, όπως η πανκυτοκερατίνη (Εικόνα 8 )και το ΕΜΑ (Epithelial Membrane Antigen), που είναι χρήσιμοι για τη διάγνωση σαρκωματόμορφου καρκινώματος, συνοβιοσαρκώματος, επιθηλιοειδούς σαρκώματος και μυοεπιθηλιώματος, η πρωτεΐνη S100 η οποία αν και δεν είναι ειδική αποτελεί πολύ χρήσιμο δείκτη για τους όγκους των ελύτρων των περιφερικών νεύρων, το μελάνωμα και το διαυγοκυτταρικό σάρκωμα, δείκτες μυϊκής διαφοροποίησης, κυρίως η δεσμίνη και η SMA, ο δείκτης CD34, για αγγειακούς όγκους, προέχον δερματοϊνοσάρκωμα, μονήρη ινώδη όγκο, στρωματικούς όγκους, ατρακτόμορφο και πλειόμορφο λίπωμα, όγκους των ελύτρων των περιφερικών νεύρων και επιθηλιοειδές σάρκωμα.[2]
![]()
Εικόνα 8 Μετάσταση σε λεμφαδένα / πανκυτοκερατίνη [2]
CD117 (Εικόνα 9 )(c-kit) για τους στρωματικούς όγκους του γαστρεντερικού συστήματος
![]()
Εικόνα 9 CD117 IHC σε GIST περιπτώσεις [3]
Οι στρωματικοί όγκοι (GISTs) είναι οι πιο συχνοί μεσεγχυματικοί όγκοι του γαστρεντερικού συστήματος (80%), μεταλλάξεις του πρωτοογκογονιδίου KIT (εξώνιο 11) στην πλειονότητα των περιπτώσεων ,υποδοχέας του KIT: δραστηριότητα τυροσινικής κινάσης, ανοσοιστοχημική ανίχνευση της KIT πρωτείνης: CD117 αντιγόνο
Πρόγνωση - Μέθοδοι ανίχνευσης
Ορμονικοί υποδοχείς (ΕR, PR, AR),: Κύριες μέθοδοι ανίχνευσης :Ενζυμικές μέθοδοι, Ανοσοϊστοχημεία. Σε καρκινώματα μαστού: ER: 24-63% των περιπτώσεων, PR: 9-37% των περιπτώσεων. Ασθενείς με θετικά επίπεδα ER και PR:έχουν μικρότερο κίνδυνο υποτροπών, καλύτερη επιβίωση [2]
· Ανοσοϊστοχημεία (προσδιορισμός της πρωτείνης)
· Φθορίζων in situ υβριδισμός (FISH) (ενίσχυση του γονιδίου)
Υπερέκφραση της HER-2/neu πρωτεΐνης σε ποσοστό 20-30% των περιπτώσεων
εύρος τιμών έκφρασης 5-85% (διαφορές στη μεθοδολογία, αξιολόγηση, χρησιμοποιηθέντα αντισώματα)
Καρκίνωμα παχέος εντέρου / E-καντχερίνη-Mεταλλάξεις του γονιδίου
Απώλεια/ελαττωμένη έκφρασή της σχετίζονται με δυσμενή πρόγνωση, αυξημένη συχνότητα υποτροπών, μικρή επιβίωση
EGF-R, 16 μελέτες (2.810 ασθενείς)
Εφαρμόσιμες τεχνικές:
ανοσοϊστοχημεία (13/16) ανοσοϊστοχημεία και Northern blot (1/16) PCR (2/16) μονοκλωνικά ή πολυκλωνικά αντισώματα, ανάλυση επιβίωσης σε 11/16 μελέτες (2.185 ασθενείς)
Η ανοσοϊστοχημική έκφραση του EGFR αποτελεί δείκτη κακής πρόγνωσης σε ασθενείς με NSCLC
Μεσεγχυματικής αρχής νεοπλάσματα
Κυτταρογενετικές μελέτες Χρωμοσωμικές ανωμαλίες - Διαμεταθέσεις γονιδίων, Διαγνωστική προσέγγιση, Προγνωστική αξία.
Συσχέτιση υπερέκφρασης της p53 πρωτεΐνης και του Ki67 (Εικόνα 10 )με επιβίωση.
![]()
Εικόνα 10 Ki67 σε συνοβιακό σάρκωμα [2]
Πρωτεΐνες κυτταρικού κύκλου (Ki67, PCNA, κυκλίνες Α, Β1, D, E, κυκλινοεξαρτώμενες κινάσες και αναστολείς τους CDK και CDKIs: p15, p16, p18, p19, p21, p27, p57)
Ογκογονίδια - Ογκοκατασταλτικά γονίδια (c-erbB-2, ras, myc, met, p53, Rb, APC, BRCA1, BRCA2)
Αυξητικοί παράγοντες (EGFR, IGFR)
Δείκτες απόπτωσης (bax, bak, bad, bcl-2, bcl-x, bcl-w)
Μόρια κυτταρικής προσκόλλησης και υποδοχείς (CD44, ιντεγκρίνες, κατενίνες, E-cadherin
Πρωτεΐνες του εξωκυτταρίου στρώματος (καθεψίνες, μεταλλοπρωτεινάσες, ενεργοποιητής του πλασμινογόνου ουροκινάση (uPA) και αναστολέας PAI-1)
Αγγειογενετικά μόρια (αγγειακός ενδοθηλιακός αυξητικός παράγοντας VEGF, b-FGF, PD-ECGF, θρομβοσπονδίνη, ενδοστατίνη, αγγειογενίνη)[2]
Η μεταγωγή σημάτων (signal transduction) είναι ο τρόπος με τον οποίο ένα κύτταρο επικοινωνεί με ένα άλλο. Η "συνομιλία" μεταξύ δύο κυττάρων περιλαμβάνει έναν μοριακό αγγελιοφόρο (ligand) από τον αποστολέα και μια περιοχή-υποδοχέα (receptor) πάνω στην μεμβράνη του κυττάρου στην οποία συνδέεται ο αγγελιοφόρος. Όταν το σήμα παραλαμβάνεται, το μήνυμα μεταφέρεται από την εξωτερική επιφάνεια του κυττάρου στον πυρήνα του κυττάρου. Τα μηνύματα μπορούν να είναι υγιή ή επιβλαβή για τον οργανισμό. Παραδείγματος χάριν, μερικά μηνύματα μπορεί να «λένε» στο κύτταρο να αυξηθεί (πράγμα που μπορεί να οδηγήσει στον καρκίνο) ή να αρχίσει η διαδικασία απόπτωσης. Μερικές από τις πιο κοινές οδούς μεταγωγής των σημάτων περιλαμβάνουν τους πρωτεϊνικούς υποδοχείς κινασών της τυροσίνης (tyrosine kinases), οι οποίες αποτελούνται από τρία βασικά μέρη:
1. Ένας εξωκυττάριος (extracellular ligand-binding domain ) υποδοχέας που βρίσκεται έξω από το κύτταρο και λαμβάνει τα εισερχόμενα σήματα
2. Μια διαμεμβρανική (transmembrane) περιοχή που διασχίζει τη μεμβράνη των κυττάρων και μεταβιβάζει τις πληροφορίες από το εξωτερικό στο εσωτερικό;
3. Μια ενδοκυτταρική περιοχή κινασών στην οποία προστίθεται ένα μόριο φωσφορικού άλατος στη τυροσίνη. Αυτή η διαδικασία είναι η αρχή ενός «καταρράκτη» αντιδράσεων στο εσωτερικό του κυττάρου και αναφέρεται ως φωσφορυλίωση "phosphorylation."[4]
Στην ογκολογία, ένα από τα σημαντικότερα δίκτυα υποδοχέων -σημάτων του τύπου των κινασών της τυροσίνης είναι μια ομάδα υποδοχέων που ανήκουν στην οικογένεια " HER ", επίσης γνωστή ως δίκτυο σημάτων ErbB. Η οικογένεια των HER υποδοχέων αποτελείται από τέσσερα κύρια μέλη καλούμενα συνήθως HER1/EGFR, HER2, HER3 και HER4. (Εικόνα 11 ) Κάθε ένας από τους υποδοχείς αυτούς εμπλέκεται στην ανάπτυξη του καρκίνου αν και ο βαθμός συμμετοχής ποικίλλει. Υπάρχει μια μη αμελητέα σχέση "αλληλεπίδρασης" μεταξύ των HER πρωτεϊνών, πράγμα που σημαίνει ότι η ενεργοποίηση ή η παρεμπόδιση της μιας μπορεί να έχει ανάλογα αποτελέσματα στις άλλες. Η κατανόηση του βιολογικού ρόλου της οικογένειάς HER έχει οδηγήσει στην ανακάλυψη και άλλων σημαντικών συστημάτων μεταγωγής σημάτων, όπως π.χ υποδοχείς που είναι διακριτοί από την οικογένειά HER αλλά έχουν τη δυνατότητα να επιδρούν με τα δικά τους σήματα στους HER –εξαρτώμενους όγκους.
![]()
Εικόνα 11 HER-2/neu (c-erbB-2)[4]
Προσδιορίζοντας την πηγή δυσλειτουργίας σε όλα τα κύτταρα, μέχρι σε ένα επίπεδο μεταγωγής σημάτων κυτταρικού πολλαπλασιασμού η διαδικασία είναι κανονική και είναι μέρος του κανονικού κύκλου πολλαπλασιασμού. Είναι η υπερέκφραση, ή η ενεργοποίηση, αυτών των σημάτων - ή η αποτυχία να αντισταθμιστούν ή να εμποδιστούν εκείνα τα σήματα -- που οδηγούν στον ανεξέλεγκτο πολλαπλασιασμό. Στην περίπτωση του γονιδίου HER2, παραδείγματος χάριν, η υπερέκφραση είναι το αποτέλεσμα μιας γενετικής αλλαγής που παράγει πολλαπλάσια αντίγραφα ενός γονιδίου το οποίο κωδικοποιεί την παραγωγή του υποδοχέα. Λόγω του πλεονάσματος των γονιδίων στο κύτταρο, δημιουργούνται υπερβολικοί αριθμοί μεμβρανικών υποδοχέων οι οποίοι όταν ενεργοποιούνται, μεταφέρουν υπερβολικά σήματα πολλαπλασιασμού που υποκινούν το κύτταρο να επιταχύνει την κυτταροδιαίρεση με τελική κατάληξη την δημιουργία όγκων. Μετάλλαξη γονιδίων HER και καρκίνος Όνομα Άλλα ονόματα Τύποι διαταραχής Είδη καρκίνου HER1 EGFR Υπερέκφραση Η μετάλλαξη οδηγεί σε ατέρμονη (non-stop) δραστηριότητα Κεφαλής, λαιμού, ουροδ.κύστης, προστάτη, νεφρού, μη μικροκυτταρικό πνεύμονα, ωοθηκών, παγκρέατος, γλοιοβλάστωμα. HER2 c-erbB-2 ErbB2 Υπερέκφραση Συν έκφραση με HER-1 βελτιώνει τη δυνατότητα πρόβλεψης για την επιθετικότητα του καρκίνου του μαστού Μαστού, ωοθηκών. HER3 ErbB3 Συνέκφραση με HER-2 βελτιώνει τη δυνατότητα πρόβλεψης για την επιθετικότητα του καρκίνου του μαστού Μαστού, παχέος, στομάχου, προστάτη, κλπ HER4 ErbB4 Συνέκφραση με HER-2 έχει προγνωστική αξία Μαστού, προστάτη, μυελοβλάστωμα (παιδιά). Ένα φυσικό χαρακτηριστικό γνώρισμα της οικογένειας των HER υποδοχέων είναι ότι το σύστημα υποδοχέων περιλαμβάνει πάντα δύο πρωτείνες-υποδοχείς σε συνδυασμό, σε έναν σχηματισμό αποκαλούμενο διμερές "dimer." Τα ομοδιμερή “homodimers” είναι συνδυασμοί δύο παρόμοιων τύπων υποδοχέων, όπως HER1/HER1 και HER3/HER3.Τα ετεροδιμερή «heterodimers» αποτελούνται από δύο διαφορετικούς υποδοχείς, όπως π.χ τα: HER1/HER2, HER1/HER3 και HER4/HER3. Τα διάφορα ετερο- και ομοδιμερή έχουν διαφορές στη ικανότητα μεταγωγής των σημάτων μέσα στο κύτταρο. Παραδείγματος χάριν, η ομο-έκφραση ορισμένων ζευγαριών (όπως HER3/HER2) είναι ισχυρότερη από άλλα όπως το HER3/HER3 ομοδιμερές, το οποίο είναι ανενεργό. Η έρευνα είναι εν εξελίξει για να γίνει καλύτερα κατανοητό γιατί διαφορετικές ενώσεις διμερών έχουν διαφορετικά αποτελέσματα και πώς αυτά τα αποτελέσματα εκδηλώνονται στο καρκινικό κύτταρο. Ογκογονίδιο HER-2/neu Το HER-2/neu είναι ένα πρωτογκογονίδιο που βρίσκεται στο χρωμόσωμα 17q11.2-12 και κωδικοποιεί ένα μέρος ενός υποδοχέα που αποτελείται από τρία μέρη: εντός κυτταροπλάσματος, επί της κυτταρικής μεμβράνης και εκτός του κυττάρου P101P. O υποδοχέας αυτός είναι μία πρωτεΐνη 185 kd πάνω στην οποία συνδέεται με τον επιδερμικό αυξητικό παράγοντα. H σύνδεση αυτή προκαλεί το σχηματισμό ομο-ετεροδιμερών που οδηγούν στην φωσφορυλίωση του ενδοκυττάριου τμήματος του υποδοχέα η οποία με τη σειρά της πυροδοτεί ένα καταρράκτη ενδοκυττάριων αντιδράσεων με τελικό αποτέλεσμα την υπερπλασία του κυττάρου. Έτσι η ενεργοποίηση του ογκογονιδίου αυτού είναι δυνατόν να οδηγήσει στην υπερβολική ή και ανεξέλεγκτη παραγωγή πρωτεϊνών-υποδοχέων οι οποίοι με τη σειρά τους ενοχοποιούνται για την προαγωγή ή ακόμα την έναρξη της διαδικασίας εξαλλαγής του κυττάρου σε νεοπλασματικό P102P. Η ενεργοποίηση του γονιδίου και η υπερπαραγωγή της πρωτεΐνης εμφανίζεται στο ένα τρίτο περίπου των όγκων στο μαστόP103P. Πάρα πολλές μελέτες έχουν γίνει για να εκτιμηθεί η προγνωστική αξία της υπερέκφρασης του HER-2/neu στους ασθενείς με καρκίνο στο μαστό. Στους ασθενείς που έχουν όγκους με θετικούς λεμφαδένες η παρουσία του HER-2/neu έχει ξεκάθαρη δυσμενή προγνωστική αξία και συνδέεται με μικρότερο διάστημα χωρίς υποτροπή της νόσου και μειωμένη επιβίωση.P104P Αντίθετα τα πράγματα είναι πιο πολύπλοκα στους ασθενείς χωρίς λεμφαδενική συμμετοχή όπου δεν έχει αποδειχτεί η προγνωστική αξία του HER-2/neu αφού τα αποτελέσματα των μελετών που έχουν γίνει μέχρι τώρα είναι αντικρουόμενα P83,93,105P. Ένα άλλο ερώτημα που προκύπτει από τη βιβλιογραφία έχει να κάνει με το κατά πόσο ανεξάρτητος είναι ως προγνωστικός δείκτης το HER-2/neu. Αν και κάποιοι ερευνητές θεωρούν το HER-2/neu ανεξάρτητο προγνωστικό δείκτη P104,105 P πολλές μελέτες έχουν δείξει ότι η υπερέκφραση του HER-2/neu συνδέεται στενά με την παρουσία και άλλων δυσμενών προγνωστικών δεικτών όπως είναι: οι διηθημένοι λεμφαδένες, το μεγάλο μέγεθος του όγκου, ο υψηλός βαθμός κακοήθειας, η απουσία οιστρογονικών και προγεστερονικών υποδοχέων, ιδιαίτερα στους όγκους με αρνητικούς λεμφαδένες κλπ. Σε μερικές από αυτές τις μελέτες το HER-2/neu χάνει την προγνωστική αξία που είχε στη univariate ανάλυση όταν γίνεται multivariate ανάλυση. Παρόλα αυτά το συμπέρασμα είναι ότι σε γενικές γραμμές η υπερέκφραση του HER-2/neu στους όγκους στο μαστό έχει δυσμενή επίδραση στη επιβίωση αυτών των ασθενών και αυτό το φαινόμενο πρέπει να εξηγηθεί σε βιολογικό και μοριακό επίπεδο P106, 107,108,109, 110P Αρκετές μελέτες έχουν προσπαθήσει να ανιχνεύσουν εάν η υπερέκφραση του HER-2/neu έχει προβλεπτική αξία ούτως ώστε να γίνει η επιλογή του καταλληλότερου χημειοθεραπευτικού ή άλλου σχήματος. Το πιο καθαρό συμπέρασμα που έχει βγει μέχρι τώρα δείχνουν ότι η υπερέκφραση του HER-2/neu συνδέεται με αυξημένη ευαισθησία στα χημειοθεραπευτικά σχήματα που περιέχουν ανθρακυκλίνες. Από την άλλη μεριά υπάρχουν κάποια δεδομένα που δείχνουν ότι το HER-2/neu συνδέεται με αυξημένη αντοχή σε άλλες ουσίες όπως είναι οι ταξάνες, η κυκλοφωσφαμίδη, η μεθοτρεξάτη, η 5-φλουοροουρακίλη και η ταμοξιφαίνη αλλά τα αποτελέσματα είναι αντικρουόμενα αφού υπάρχουν και μελέτες που δείχνουν ευαισθησία σε αυτές τις ουσίες P105,106,107,111P Η διαφαινόμενη σημασία του HER-2/neu στη πρόγνωση και στη θεραπεία των ασθενών με καρκίνο του μαστού το έκανε το ίδιο θεραπευτικό στόχο. Έτσι αναπτύχθηκε μετά από πειράματα στο εργαστήριο ένα μονοκλωνικό ειδικό αντίσωμα που λέγεται Herceptin (trastuzumab) και χορηγείται στους ασθενείς στοχεύοντας το ίδιο το HER-2/neu στα καρκινικά κύτταρα P112P. Mάλιστα το αντίσωμα αυτό έχει δείξει πολύ καλά αποτελέσματα ακόμα και όταν χορηγείται μόνο του σε ασθενείς στους οποίους δεν απέδωσε η χημειοθεραπεία P113P. Πρόσφατα ιδιαίτερο ενδιαφέρονP114P έχει αποκτήσει η εμπλοκή της ενεργοποίησης του γονιδίου αυτού μαζί με άλλα στων αρχική φάση της ογκογενετικής διαδικασίας στο μαστό η οποία μέχρι τώρα αποδιδόταν αποκλειστικά στην μεσολάβηση άμεσα η έμμεσα των οιστρογόνων και των υποδοχέων τους.[4]
Συνδυασμένη έκφραση των p53, Bcl-2, και p21WAF-1 πρωτεϊνών στον καρκίνο των πνευμόνων και σε προκαρκινοματώδεις αλλοιώσεις: σύνδεση με τα κλινικά χαρακτηριστικά
Εξετάστηκαν η πρωτεϊνική έκφραση των p53, p21WAF-1, και Bcl-2 (Εικόνα 12 )σε κακοήθη και καλοήθη βρογχικά δείγματα αποκτηθέντα κατά τη διάρκεια βρογχοσκόπησης από 60 ασθενείς με καρκίνο πνευμόνων. Είκοσι έξι (43.3%), 36 (60%), και 20 (33.3%) από τους όγκους ήταν p53, p21WAF-1, και Bcl-2 θετικό, αντίστοιχα. Υψηλά επίπεδα p53 και Bcl-2 έκφρασης προσδιορίστηκαν σε προχωρημένες προκαρκινοματώδεις, αλλοιώσεις ενώ υπερπλασίες, πλακώδεις μεταπλασίες, και ήπιες δυσπλασίες έδειξαν χαμηλά επίπεδα έκφρασης. Δεν υπήρξε καμία διαφορά μεταξύ πρώϊμων και προχωρημένων προκαρκινοματωδών αλλοιώσεων στο επίπεδο της έκφρασης των p21WAF-1. Ένα ιστορικό βαριού καπνίσματος συνδέθηκε με την έκφραση των p21WAF-1, στις προκαρκινοματώδεις, αλλοιώσεις (p = 0.022) και όγκους (p = 0.032). p53(-)/p21WAF-1(++)/bcl-2(-)ήταν ο μόνος σημαντικός ανεξάρτητος προάγγελος από χαμηλότερη κλινική κατάσταση(OR: 0.88, p = 0.038). Στη μεταβλητή ανάλυση, επιβίωσης NSCLC οι ασθενείς επηρεάστηκαν από την κατάσταση της ασθένειας (p <0.001) και ιστολογία όγκων (p = 0.018). Ενώ η ενιαία-πρωτεϊνική έκφραση δεν συνδέθηκε με την πρόγνωση, συνδυασμένος ανοσοφαινότυπος p53(-)/p21WAF-1(++)/bcl-2(-) προβλέπει πιό μακροχρόνια επιβίωση (p = 0.03). Στην πολλών μεταβλητών ανάλυση, μόνο TNM κατάσταση βρέθηκε να είναι ένας προγνωστικός παράγοντας για NSCLC. Ολοκληρώνοντας αυτές οι αλλαγές των p53 και Bcl-2 μπορούν να συμβούν νωρίς στη βρογχική καρκινογένεση και η απουσία από αυτές τις αλλαγές σε συνδυασμό με την υπερέκφραση των p21WAF-1, μπορέστε να συνδεθεί με συμπεριφορά λιγότερο επιθετικού όγκου. Αυτό είναι μια από τις πολλές κλινικές μελέτες που επιδιώκουν να προσδιορίσουν μοριακούς δείκτες από τα διάφορα στάδια του καρκίνου των πνευμόνων. Η παρούσα μελέτη προσθέτει κάποια στοιχεία για εκείνους τους μοριακούς δείκτες που μπορούν να είναι προφητικοί για τη βιολογία και την επιθετικότητα όγκων και μπορεί να συμπληρώσει το TNM στάδιο για προγνωστικούς λόγους.[5]
![]()
Εικόνα 12 bcl-2 σε αδενοκαρκίνωμα μαστού[5]
Ο ενεργοποιητής του πλασμινογόνου (UPA). Ένας νέος δείκτης για τον καρκίνο του παχέος εντέρου
Η σταδιοποίηση του καρκίνου του παχέος εντέρου κατά DUKES' αποτελεί μέχρι σήμερα τον καλύτερο δείκτη πρόγνωσης αν και υπάρχει μεγάλη ανάγκη για περαιτέρω υποδιαίρεση, κυρίως των σταδίων Β και C, σε περισσότερες κατηγορίες που δίνουν τη δυνατότητα ιδιαίτερης θεραπευτικής προσέγγισης. Αυτό ίσως γίνει εφικτό με τη διερεύνηση νέων βιολογικών δεικτών. Η διήθηση και η νετάσταση του καρκίνου διευκολύνονται από αρκετά πρωτεολυτικά ενζυμικά συστήματα μεταξύ των οποίων και οι ενεργοποιητές πλασμινογόνου. Μελετήθηκε η έκφραση του ενεργοποιητή του πλασμινογόνου της ουροκινάσης (UΡΑ) σε 64 ζεύγη κυτοσολίων ιστών παχέος εντέρου (καρκινικό/μη καρκινικό των ίδιων ασθενών) με τη μέθοδο ELISA, καθώς επίσης με ανοσοϊστοχημική μέθοδο σε τομές παραφίνης των όγκων των ίδιων ασθενών. για τη στατιστική μελέτη χρησιμοποιήθηκαν οι δοκιμασίες ΜANN-WHITNEY, KRUSKAL-WALLIS, X2 ΚΑΙ FISHERS EXACT. Εξήντα δύο ασθενείς είχαν υψηλότερη συγκέντρωση UΡΑ (96,8%) στον καρκινικό απ' ό,τι στον μη καρκινικό ιστό, ενώ δύο είχαν χαμηλότερη στον καρκινικό (3,2%) απ' ό,τι στον μη καρκινικό. Οι συγκεντρώσεις του UΡΑ κυμαίνονταν μεταξύ 0,30 και 2,80 NG/MG πρωτεϊνης, με μέσο όρο±μέση απόκλιση 1,92±0,47 NG/MG πρωτεϊνης για τον καρκινικό ιστό ενω για τον μη καρκινικό κυμαίνονταν μεταξυ 0,18 και 0,75 με μέσο όρο±σταθερή απόκλιση 0,29±0,11 NG/MG πρωτεϊνης. Στατιστική ανάλυση έδειξε ότι η διαφορά της συγκέντρωσης του UΡΑ μεταξύ καρκινικών και μη καρκινικών ιστών παχέος εντέρου των ίδιων ασθενών είναι στατιστικά σημαντική (Ρ<0,001). Επίσης στατιστική ανάλυση έδειξε θετική σχέση μεταξύ της συγκέντρωσης του UΡΑ και του βαθμού διαφοροποίησης (GRADE) (Ρ<0,001), καθώς επίσης και της διήθησης των λεμφαδένων (Ρ=0,045), όχι όμως και του σταδίου κατά DUKES'. Μελέτη της έκφρασης του UΡΑ με ανοσοϊστοχημεία έδειξε θετική σχέση με το GRADE, το στάδιο κατά DUKES' και τη διήθηση των λεμφαδένων (Ρ<0,001, Ρ<0,001, Ρ=0,002 αντίστοιχα). Μελέτη μεγαλύτερου αριθμού δειγμάτων διεξάγεται στο εργαστήριο με στόχο την διερεύνηση της πιθανότητας χαρακτηρισμού του UΡΑ ως νέου προγνωστικού δεικτη για τον καρκίνο του παχέος εντέρου.[6]
Καρκίνος ουροδόχου κύστηςΟ καρκίνος της ουροδόχου κύστης αντιπροσωπεύει τον 4ο συχνότερο καρκίνο στους άνδρες και τον 10ο στις γυναίκες, με ποσοστό υποτροπής 70% και 30% των υποτροπών να εξελίσσονται. Περίπου 60% των ασθενών με προχωρημένο καρκίνο ουροδόχου κύστης θα έχει αυξημένα επίπεδα ορού, τουλάχιστον ενός από τους παρακάτω καρκινικούς δείκτες: βHCG, CEA, CA125 και CA19.9, οι οποίοι αποτελούν και δείκτες ανταπόκρισης στην χημειοθεραπεία. Χρησιμοποιούνται ήδη στην κλινική πράξη οι παρακάτω διαγνωστικές εξετάσεις καρκινικών δεικτών των ούρων για τον καρκίνο της ουροδόχου κύστης: το ΒΤΑstat test (ποιοτική ανοσοχρωματογραφική εξέταση που ανιχνεύει το ειδικό αντιγόνο hCFHrp), το NMP22 test (ανοσολογική εξέταση ποσοτικής ανίχνευσης της πυρηνικής δομικής πρωτεΐνης 22), το UBC test (ανοσολογική ποιοτική αλλά και ποσοτική μέτρηση των επιπέδων τμημάτων των κυτταροκερατινών 8 και 18), το GYFRA-21 test (ανοσοενζυμική μέτρηση κυτταροκερατίνης 21-1) και το Aura Tek FDP test (μέτρηση προϊόντων καταβολισμού ινώδους/ινωδογόνου). Η κυτταρομετρία ροής μελετώντας την πλοειδικότητα του DNA και τις φάσεις πολαπλασιασμού υπερέχει της κυτταρολογικής εξέτασης των ούρων στην διάγνωση του καρκίνου της κύστης. Χρησιμοποιούνται επίσης και οι τεχνικές γονιδιακής υβριδοποίησης (CGH) και κυτταρικής μορφομετρίας (CIA). Υπό μελέτη είναι οι παρακάτω δείκτες: οι ρυθμιστικές πρωτεΐνες του κυτταρικού κύκλου όπως η τελομεράση και ο αναστολέας των κινασών κυκλίνης p21Cip1, τα αντιγόνα ομάδων αίματος (ΑBH, Lewis και Thomsen-Friedenreich), οι δείκτες πολλαπλασιασμού (π.χ., Ki67), ογκογονίδια (π.χ., ras, bcl-2, c-Erb-B2), τα ογκοκατασταλτικά γονίδια (π.χ., p53 και Rb), οι παράγοντες αγγειογένεσης και μετάστασης (π.χ., VEGF, VPF, PDECGF, MMPs, ΗΑ), οι αυξητικοί παράγοντες και οι υποδοχείς τους (π.χ., EGF και EGFR, (Εικόνα 13 ) TGF-a), τα μόρια συγκόλλησης (π.χ., Ε-cadherin) και άλλα.[7]
Εικόνα 13 EGF-R σε αδενοκαρκίνωμα [2] Καρκίνος νεφρούΗ επίπτωση του καρκίνου του νεφρού έχει αυξηθεί την τελευταία εικοσαετία. Eνοχοποιούνται γονίδια όπως το ογκογονίδιο c-met και το von Hippel-Lindau (VHL) γονίδιο που κωδικοποιεί μεταγραφικό παράγοντα επεξεργασίας του RNA. Έχει διαπιστωθεί ότι το CEA είναι αυξημένο στις μισές περίπου περιπτώσεις. Άλλοι δυνητικοί καρκινικοί δείκτες είναι το CA50 και TPA. Υπό μελέτη μοριακοί καρκινικοί δείκτες είναι διάφοροι παράγοντες απόπτωσης και αγγειογένεσης καθώς και δείκτες πολλαπλασιασμού. Τέλος, η καρυοτυπική ανάλυση με in situ υβριδισμό και η RFLP ανάλυση του γενετικού υλικού, βοηθά σημαντικά στη γενετική ταξινόμηση των νεοπλασμάτων του νεφρού.[7] Καρκίνος προστάτηO καρκίνος του προστάτη αποτελεί την δεύτερη συχνότερη αιτία θανάτου από καρκίνο στους άνδρες, μετά από τον καρκίνο του πνεύμονα, με το μειονέκτημα σε πάνω από το ήμισυ των περιπτώσεων ο καρκίνος να διαγνώσκεται σε προχωρημένο στάδιο. Η εισαγωγή της χρήσης του ειδικού προστατικού αντγόνου (PSA), (Εικόνα 14 )την τελευταία εικοσαετία, προσέθεσε τον πιο σημαντικό καρκινικό δείκτη για τον καρκίνο του προστάτη, στις χρησιμοποιούμενες εξετάσεις του προστατικού κλάσματος της όξινης φωσφατάσης και της ΑLP. Προκειμένου να αυξηθεί η ειδικότητα και ευαισθησία του PSA, έχουν προταθεί παράμετροι όπως το κλάσμα ελεύθερου προς ολικό PSA, η ταχύτητα αύξησης του PSA και η πυκνότητα του PSA και η σχέση του PSA με την ηλικία. Η χρησιμότητα του PSA δεν περιορίζεται μόνο στην πρώιμη διάγνωση και ριζική αντιμετώπιση του καρκίνου του προστάτη, αλλά και στον έλεγχο της αποτελεσματικότητας της χειρουργικής αντιμετώπισης, ορμονοθεραπείας, χημειοθεραπείας και ακτινοθεραπείας, αποτελώντας παράλληλα έναν πολύ χρήσιμο προγνωστικό δείκτη.[7]
Εικόνα 14 PSA σε αδενοκαρκίνωμα προστάτη [2] Υπό μελέτη είναι οι παρακάτω δείκτες που παίζουν ρόλο και στην ανάπτυξη ανδρογόνο-ανεξαρτησίας και αποτυχίας του ανδρογονικού αποκλεισμού: οι αυξητικοί παράγοντες με τους αντίστοιχους υποδοχείς (π.χ., EGF, KGF, IGF-I, IGF-IΙ), οι αποπτωτικές και αντιαποπτωτικές πρωτεΐνες (π.χ., Βax, Bcl-2, Bcl-X, Mcl-1), οι κυκλίνες (π.χ., D, E, Α), οι κινάσες των κυκλινών και οι αναστολείς τους (π.χ., CDK4, CDK6, CDK2, p27Kip1, p15INK4B, p16INK4A), (Εικόνα 15) οι τελομεράσες, τα προϊόντα ογκογονιδίων και ογκοκατασταλτικών γονιδίων (π.χ., ΗΕR-2/Neu, ras, c-met, p53, Rb), οι παράγοντες αγγειογένεσης και επέκτασης (VEGF, C-CAMI, MUC1, MUC2, MMPs, TIMPs, uPA) οι δείκτες νευροενδοκρινικής διαφοροποίησης (π.χ., NGF) και άλλα.[7]
Εικόνα 15 p27 σε καρκίνωμα [2] Στον εντοπισμό των παραπάνω δεικτών βοηθούν και οι διάφορες σύγχρονες τεχνικές ανίχνευσης γενετικών διαταραχών, όπως στην περίπτωση των χρωμοσωμιακών μεταλλάξεων, των πολυμορφισμών (π.χ., των υποδοχέων των ανδρογόνων; AR, ή της βιταμίνης D; VDR), της γονιδιακής ενίσχυσης (π.χ., του AR) και της απώλειας της ετεροζυγωτίας (LOH). Τα ειδικά αντιγόνα του προστάτη (π.χ., PSA, PSMA, PSCA, PCGEM1, DD3, PSGR) δεν αποτελούν μόνο ειδικούς δείκτες, αλλά και βοηθούν στην ειδικότητα της γονιδιακής θεραπείας (αποκατάστασης ή αυτοκτονίας) αλλά και στην αποτελεσματικότητα της ανοσοθεραπείας.[7] Καρκίνος όρχεωνΟ καρκίνος των όρχεων είναι ο πιο συχνός καρκίνος στους νεαρούς άρρενες με επίπτωση που έχει διπλασιαστεί τα τελευταία 25 έτη. Ενοχοποιούνται διαταραχές ογκογονιδίων και αυξητικών παραγόντων: c-kit, hst1, c-myc, κυκλίνης D και PTHLH. Καθιερωμένοι καρκινικοί δείκτες είναι η AFP και η βHCG. Επίσης χρησιμοποιούνται με διαφορετικοί ειδικότητα και ευαισθησία στους διάφορους ιστολογικούς τύπους οι παρακάτω δείκτες: CEA, LDH, γ-GT, PLAT, SSP1, τεστοστερόνη, ειδική β1-γλυκοπρωτείνη της κύησης, πλακουντιακή ALP και άλλα. [7] |
Εισαγωγή
Ως καρκινικοί δείκτες ορίζονται ουσίες που έχουν τη δυνατότητα να προσδιορισθούν ποσοτικά και να παράσχουν πληροφορίες σχετικά με την ύπαρξη, την φύση και την ανάπτυξη του όγκου, την μεταστατική του εξάπλωση και την ανταπόκριση ή όχι στην χημειοθεραπεία (ΧΜΘ).
Η επιλογή της θεραπείας στηρίζεται κυρίως 1) στην ύπαρξη προγνωστικών παραγόντων όπως το ΤΝΜ σύστημα, 2) στην έκταση των λεμφαδενικών (Ν) μεταστάσεων και 3) στην γενική κατάσταση του ασθενούς.
Πολλές φορές οι ασχολούμενοι με τον καρκίνο του πνεύμονα έρχονται αντιμέτωποι με ασθενείς που παρά την διαφορετική τους πρόγνωση, καταλήγουν σύντομα κι αυτό δείχνει ότι άλλα, ειδικά χαρακτηριστικά του όγκου παίζουν ρόλο στον καθορισμό της κλινικής πορείας του ασθενούς. Η γνώση των βιολογικών μεταβολών που συνοδεύουν τον καρκίνο του πνεύμονα είναι απαραίτητη προκειμένου να κατανοήσουμε τους γενετικούς και κυτταρικούς μηχανισμούς που είναι υπεύθυνοι για την ανάπτυξη και τον έλεγχο αύξησης του όγκου.
Η κλινική αξία της χρήσης των καρκινικών δεικτών μέχρι σήμερα περιορίζεται α) στην προγνωστική αξιολόγηση και β) στην παρακολούθηση της πορείας της νόσου, είτε είναι χειρουργήσιμη είτε όχι.
Στόχος της έρευνας σε αυτό το πεδίο είναι να βρεθούν ουσίες με υψηλή προγνωστική αξία στη διάγνωση του καρκίνου του πνεύμονα που θα καθιστούν ικανό τον κλινικό γιατρό να προγραμματίζει με μεγαλύτερη ακρίβεια τους θεραπευτικούς του χειρισμούς, να αξιολογεί την ανταπόκριση στη ΧΜΘ και να προσδιορίζει την χρονικότητα των υποτροπών.
Οι βιολογικοί, μοριακοί και γενετικοί δείκτες που αναζητούνται στον καρκίνο του πνεύμονα είναι:
* Καρκινοεμβρυϊκό αντιγόνο (CEA)
* Καρκινικό αντιγόνο 15-3 (CA 15-3)
* Κυτταροκερατίνη 21-1 (CYFRA 21-1)
* Νευροενδοκρινική ενολάση (NSE)
* Β-Β Κρεατινική κινάση (Β-Β creatine kinase)
* Αντιγόνο πλακώδους καρκινώματος (SCC)
* Ιστικό πολυπεπτιδικό αντιγόνο (TPA)
* Ειδικό Ιστικό Πολυπεπτιδικό Αντιγόνο (TPS).
Νεότεροι δείκτες έχουν χρησιμοποιηθεί στον καρκίνο του πνεύμονα κι αυτοί είναι οι:
* Υποδοχείς ορμονικών υποδοχέων στεροειδικών ορμονών, καθώς και μοριακοί και γενετικοί καρκινικοί δείκτες όπως:
* Ογκογονίδια και ογκοπρωτεϊνες
* Ογκοκατασταλτικά γονίδια και πρωτεϊνες
* Αυξητικοί παράγοντες του όγκου
* Παραγωγή χειμερικών πρωτεϊνών
* Απώλεια ετεροζυγωτείας και μικροδορυφορικό DNA
* Προσδιορισμός Τελομεράσης.
Καρκίνος πνεύμονα
Ο καρκίνος του πνεύμονα είναι ο πιό κοινός και θανατηφόρος καρκίνος παγκοσμίως. Περίπου το 80% του καρκίνου του πνεύμονα αποτελεί ο ΜΜΚΠ που σταδιοποιείται κατά ΤΝΜ σύμφωνα με την πρόταση των C.Mountain, Ts. Naruke.
Η βασική θεραπευτική αντιμετώπιση για τα πρώϊμα στάδια του ΜΜΚΠ (Ι, ΙΙ, ΙΙΙα) που αποτελούν το 20-25% των καρκίνων, είναι η χειρουργική εξαίρεση. Σε αυτά τα στάδια, η αύξηση των επιπέδων των καρκινικών δεικτών αποτελεί ένδειξη φτωχής πρόγνωσης (διάστημα ελεύθερο νόσου και συνολική επιβίωση).
Η αξιολόγηση των καρκινικών δεικτών προσδιορίζει περιπτώσεις υψηλού κινδύνου για υποτροπή και ασθενείς που μπορούν να δεχθούν ΧΜΘ ανοσοενισχυτικός και nνεο- ανοσοενισχυτικός.
Ο μετεγχειρητικός προσδιορισμός των καρκινικών δεικτών χρησιμεύει:
* Στην εκτίμηση της υπολειμματικής νόσου
* Στον πρώϊμο προσδιορισμό των υποτροπών
Οι συνήθως χρησιμοποιούμενοι καρκινικοί δείκτες στον ΜΜΚΠ φαίνονται στον πίνακα.
Όνομα | Σύντμηση | Βιοχημικές ιδιότητες |
Καρκινοεμβρυικό αντιγόνο | CEA | Γλυκοπρωτεϊνη |
Καρκινικό αντιγόνο 125 | CA 125 | Βλεννώδης ουσία προσδιορισμένη με μονοκλωνικά αντισώματα |
Αντιγόνο πλακώδους Καρκινώματος | SCC-Ag | Γλυκοπρωτεϊνικό κλάσμα καρκινικού αντιγόνου Τ4 |
Ιστικό πολυπεπτιδικό αντιγόνο | TPA | Τμήμα των κυτταροκερατινοκινών 8,18,19 |
Κυτταροκερατίνη 21-1 | CYFRA 21-1 | Τμήμα κυτταροκερατίνης 19 |
Ειδικό ιστικό πολυπεπτιδικό αντιγόνο | TPS | Τμήμα κυτταροκερατίνης 18 |
Συμπερασματικά πρέπει να πουμε ότι οι καρκινικοί δείκτες μπορεί να είναι χρήσιμοι στον ΜΜΚΠ σε 4 περιπτώσεις:
1. Ως προγνωστικοί δείκτες της έκτασης της νόσου προεγχειρητικά σε ασθενείς με εντοπισμένη νόσο. Σε απουσία απομακρυσμένων μεταστάσεων, ίσως είναι υποψήφιοι για νεο-ανοσοενισχυτική ή ανοσοενισχυτική θεραπεία.
2. Στην λήψη απόφασης για έναρξη ΧΜΘ σε ασθενείς με εκτεταμένη νόσο. Οι καρκινικοί δείκτες θα μπορούσαν να βελτιώσουν την ικανότητά μας να επιλέξουμε τους κατάλληλους ασθενείς και ίσως να μας βοηθήσουν στην επιλογή των ΧΜΘ φαρμάκων.
3. Στην μετεγχειρητική παρακολούθηση ασθενών. Οι καρκινικοί δείκτες πρέπει να αποτελούν μέρος των θεραπευτικών χειρισμών και η χρησιμότητά τους πρέπει να συγκρίνεται με συγκεκριμένους διαγνωστικούς χειρισμούς κατά τον προσδιορισμό των υποτροπών.
4. Στην παρακολούθηση της απάντησης στην θεραπεία γεγονός που θα μας βοηθήσει να αλλάξουμε θεραπευτική προσέγγιση αν ο ασθενής δεν απαντά περισσότερο στην εφαρμοζόμενη θεραπεία.
Νευροενδοκρινικοί δείκτες
Σε περισσότερο από 80% ΜΚΠ έχουν προσδιοριστεί Νευροενδοκρινικοί δείκτες με κυριο εκπρόσωπο την NSE (Νευροενδοκρινική Ειδική Ενολάση). Νευροενδοκρινική δραστηριότητα παρουσιάζει και ο ΜΜΚΠ (23%) και η NSE συσχετίζεαι με καλλίτερη απάντηση στη ΧΜΘ.
Σε εργασία των NE Kushlinsky αναφέρεται ότι η NSE παρουσιάζει διαγνωστική ευαισθησία 76% για τον ΜΚΠ με ψηλότερες τιμές σε εκτεταμένη νόσο.
Η αυξηση της NSE σε ΜΜΚΠ παρουσιάζει διαφορές στο πλακώδες (34%) και στο αδενοκαρκίνωμα (43,8%). Συμφωνα με τον Rastel η NSE δεν έχει την απαιτουμενη ειδικότητα και ευαισθησία προκειμένου να χρησιμοποιηθεί στην δ/δ του ΜΚΠ από τον ΜΜΚΠ. Η μέτρησή της μπορεί να χρησιμευσει σην παρακολουθηση της πορείας του ΜΚΠ.
Μοριακοί και γενετικοί δείκες
Ογκογονίδια: Σ' αυτά ανήκουν κυρίως οι οικογένειες Myc και Ras.
Στην οικογένεια Ras συγκαταλέγονται τα HRAS1, KRAS2, NRAS και ο KRAS2 παρουσιάζει μετάλλαξη στη θέση 12. Απαντάται 90% στο Αδενοκαρκίνωμα και συσχετίζεται με αυξηση σο ρυθμό μεγένθυνσης του όγκου και μικρότερη επιβίωση.
Η οικογένεια Myc περιλαμβάνει τα c- Myc, N- Myc, L- Myc. Παρατηρείαι μεγένθυνση ή υπερέκφραση κυρίως στο c- Myc. Αυτή η βλάβη απαντάται 80-90% σε ΜΚΠ και 10% σε ΜΜΚΠ και δηλώνει τάση για υποτροπή μετά ΧΜΘ καθώς και κακή πρόγνωση.
Ογκοκατασταλτικά γονίδια: Παρατηρείται απώλεια λειτουργικότητας. Χαρακτηριστικοί εκπρόσωποι είναι το γονίδιο του Ρετινοβλαστώματος που απαντάται κυρίως στο ΜΚΠ και λιγότερο σχνά σε ΜΜΚΠ, το γονίδιο p53 (Εικόνα 16 )που παρατηρείται 75-78% σε ΜΚΠ και 50% σε ΜΜΚΠ. Οι μεταλλάξεις αυτές σχετίζονται με μεταστάσεις σε πυλαίους και μεσοθωρακικους λεμφαδένες. Οι απώλειες στο χρωμόσωμα 3ρ παρατηρουνται 100% στο ΜΚΠ και 50% στο ΜΜΚΠκαι αφορουν σε ομόζυγη εξάλειψη των περιοχών 3ρ12-3ρ22.
![]()
Εικόνα 16 p53 σε ινοσάρκωμα [2]
Αλλοι παράγοντες που ο ρόλος τους ακόμη συζητείται σην κλινική αξιολόγηση των καρκίνων του πνευμονα είναι οι αυξητικοί παράγονες GRP, EGF, GF-a, IGF-1 και ο προσδιορισμός επιφανειακών επιτόπων που αναμένεται να πλουτίσον τις γνώσεις μας και να συμβάλλον στην πληρέστερη αντιμετώπιση το καρκινοπαθή.[8]
Θεραπεία
Η κλινική έρευνα της θεραπευτικής των κακοήθων νεοπλασμάτων επικεντρώνεται στους διαρκώς αναγνωριζόμενους μοριακούς δείκτες που εμπλέκονται στην παθογένειά τους.
Την τελευταία δεκαετία εξελίχθηκε η προσπάθεια ανάπτυξης θεραπευτικών παραγόντων με ικανότητα στόχευσης και αναστολής της λειτουργίας αυτών των δεικτών.[2]
1846 πρωτείνη Bence-Jones
1940 όξινη φωσφατάση
1963 α-φετοπρωτείνη (AFP)
1965 καρκινοεμβρυικό αντιγόνο (CEA)
1975 μονοκλωνικά αντισώματα
1980 CA 125, CA 15.3, CA 19.9 ειδικό προστατικό αντιγόνο (PSA)
1970-1980 ογκογονίδια, ογκοκατασταλτικά γονίδια
2001 PCR, μικροσυστοιχίες, πρωτεομική, σπεκτρομετρία μάζας [2]
· Σάρωση γενικού πληθυσμού
· Διάγνωση και διαφορική διάγνωση συμπτωματικών ασθενών
· Σταδιοποίηση της νόσου
· Εκτίμηση της πρόγνωσης
· Εκτίμηση ύφεσης ή υποτροπής της νόσου
· Εκτίμηση ανταπόκρισης στη θεραπεία
1. Screening γενικού πληθυσμού.
· περιορισμένη
· μικρή ευαισθησία (οι περισσότεροι δείκτες ανιχνεύονται σε υψηλά επίπεδα σε προχωρημένα στάδια)
· μικρή ειδικότητα
· έκφραση σε ποικιλία νεοπλασμάτων και σε μη νεοπλασματικές παθήσεις
2. Διάγνωση και διαφορική διάγνωση συμπτωματικών ασθενών
· περιορισμένη
· μικρή ευαισθησία
· μικρή ειδικότητα
για επιλεγμένες ομάδες ασθενών, π.χ υψηλού κινδύνου, η συστηματική ανάλυση δεικτών πιθανώς να οδηγήσει σε λεπτομερέστερο έλεγχο (ενδοσκόπηση, απεικονιστικές μέθοδοι)
3. Σταδιοποίηση της νόσου.
· περιορισμένη
4. Εκτίμηση πρόγνωσης για την εξέλιξη της νόσου.
· σχετικά περιορισμένη
· κάποιοι δείκτες φαίνεται να έχουν προγνωστική αξία
5. Εκτίμηση ανταπόκρισης στη θεραπεία.
· σημαντική
· λίγοι δείκτες: καρκίνωμα μαστού ER, PR, c-erbB2 (Εικόνα 17 )
![]()
![]()
Εικόνα 17 [2]
6. Εκτίμηση υποτροπής ή ύφεσης της νόσου.
· περιορισμένη
· υποτροπή νόσου χωρίς αύξηση τιμών
· μη ειδική αύξηση χωρίς επιδείνωση της νόσου
διακοπή του θεραπευτικού σχήματος ή υπερθεραπεία [2]
· Γονίδια και κωδικοποιούμενες πρωτείνες
· Κυτταρική εντόπιση των πρωτεινών
· Δομή και λειτουργία των πρωτεινών
· Βιοχημικά μονοπάτια
· Φάσμα μεταλλάξεων
· Τεχνολογίες (γενομική, πρωτεομική)
Τα νοσήματα θα ταξινομούνται με βάση μοριακή και όχι μορφολογική ανάλυση.
Η έγκαιρη διάγνωση θα είναι δυνατή με την αναζήτηση συγκεκριμένων γονιδιακών ή πρωτεινικών profiles, που θα αποτελούνται από πολλούς μοριακούς δείκτες (biomarkers).
Οι μοριακοί δείκτες θα αποτελούν στόχους θεραπευτικών στρατηγικών.[2]
Οι συστοιχίες µικροκηλίδων ολιγονουκλεοτιδίων παρέχουν ένα σηµαντικό εργαλείο για την ταυτόχρονη ανίχνευση πολλαπλών µεταλλάξεων σε γονίδια.
Πλεονεκτούν έναντι των κλασσικών µεθόδων ανάλυσης του DNAως προς την ταχύτητα, την απλότητα και το κόστος της ανάλυσης.
Απαιτείται προσεκτική ρύθµιση των σύνθηκών ακινητοποίησης των ολιγονουκλεοτιδίων ανιχνευτών καθώς και των συνθηκών υβριδισµού και έκπλυσης ώστε να επιτευχθεί επαρκές σήµα φθορισµού και υψηλή ειδικότητα.
Είναι ιδανικές για µελέτες σε επίπεδο πλυθησµού, για πρόγνωση ασθενειών, ανακάλυψη νέων πολυµορφισµών που µπορεί να σχετίζονται µε έναν συγκεκριµένο γονότυπο.[9]
Στον μη-μικροκυτταρικό καρκίνο του πνεύμονα (Non-Small Cell Lung Cancer, NSCLC) έχουν βρεθεί μεταλλάξεις του EGFR. Ασθενείς φορείς των μεταλλάξεων αυτών ανταποκρίνονται σε στοχευμένη θεραπεία με αναστολείς του EGFR (gefitinib). Οι μεταλλάξεις αυτές (σημειωμένες με κόκκινο στο σχήμα) εντοπίζονται στην περιοχή του γονιδίου (εξώνια 18-21) που αντιστοιχεί στην περιοχή κινάσης τυροσίνης. (Εικόνα 18 )
Η διαδικασία της ανάλυσης των μεταλλάξεων του EGFR περιλαμβάνει τα εξής στάδια:
1. απομόνωση DNA από κύτταρα του όγκου
2. εκλεκτική ενίσχυση των εξωνίων 18-21 με PCR
3. προσδιορισμός της αλληλουχίας των βάσεων και σύγκριση με αλληλουχίες αναφοράς. [10]
 Εικόνα 18 [10]
Εικόνα 18 [10]
Ο γενετικός έλεγχος των συμπαγών όγκων προσφέρει σημαντικά δεδομένα χρήσιμα στη διάγνωση, στην πρόγνωση και στην επιλογή της κατάλληλης θεραπείας [11, 12]. Οι αναλύσεις με την κλινικά σημαντικότερη αξία εστιάζονται κυρίως στη διαφορική διάγνωση των μπλε μικροστρογγυλοκυτταρικών όγκων. Στα νεοπλάσματα αυτά η γενετική ανάλυση αποσαφηνίζει κατηγορηματικά τη διάγνωση με τον εντοπισμό ειδικών μεταθέσεων για το κάθε είδος όγκου. Η μεθοδολογία έχει επεκταθεί και σε ορισμένα άλλα είδη μεσεγχυματογενών όγκων [12]
Σήμερα διεξάγονται οι ακόλουθες αναλύσεις:
Γενετικές αναλύσεις συμπαγών όγκων με την τεχνική RΤ-PCR
| Χρωμοσωμική Ανωμαλία | Γονιδιακή Ανακατάταξη |
Σάρκωμα Ewing (ES) | t(11;22) | EWS/FLI1 |
Σάρκωμα Ewing (ES) | t(21;22) | EWS/ERG |
Συνοβιοσάρκωμα ( SS ) | t(X;18) | SYT/SSX |
Ραβδομυοσάρκωμα ( RH ) | t(2;13) | PAX3/FKHR |
Ραβδομυοσάρκωμα ( RH ) | t (1;13) | PAX7/FKHR |
Δεσμοπλαστικός μικροστρογγυλοκυτταρικός όγκος( D ) | t(11;22) | EWS/WT1 |
Μυξοειδές Λιποσάρκωμα ( ML ) | t(12;16) | FUS/CHOP |
Μυξοειδές Λιποσάρκωμα ( ML ) | t(12;22) | EWS/CHOP |
Σάρκωμα εκ διαυγών κυττάρων ( CCS ) | t(12;22) | EWS/ATF1
|
Οι αναλύσεις για ML πραμγατοποιούνται με την τεχνική long-PCR.
Επεξηγήσεις: RT-PCR: reverse transcription-PCR, t: μετάθεση.
Ο γενετικός έλεγχος ασθενών με Χρόνια Μυελογενή Λευχαιμία (CML), Οξεία Μυελογενή Λευχαιμία (AML) και Οξεία Λεμφοβλαστική Λευχαιμία (ALL) αποτελεί εργαλείο για τη διάγνωση, την πρόγνωση και την παρακολούθηση αυτών των ασθενειών. Επιπλέον, ο γενετικός έλεγχος καθιερώνεται σταδιακά ως εξέταση ρουτίνας στα Μυελοδυσπλαστικά Σύνδρομα καθώς και στη Χρόνια Λεμφοκυτταρική Λευχαιμία (CLL), ενώ σύντομα αναμένεται να επεκταθεί και στα Μυελώματα, δεδομένου ότι νέοι γενετικοί δείκτες ταυτοποιούνται διεθνώς [13, 14].
Η πρώτη χρωμοσωμική ανωμαλία που συνδέθηκε με τη νεοπλασία, το χρωμόσωμα Φιλαδέλφεια, εντοπίστηκε σε κακοήθη αιματολογική νόσο [15]. Έκτοτε, στους διάφορους τύπους λευχαιμιών και λεμφωμάτων έχουν ανιχνευθεί περίπου 200 χρωμοσωμικές μεταθέσεις [16].
Οι περισσότερες είναι ειδικές για συγκεκριμένους μορφολογικούς, ανοσοφαινοτυπικούς και ιστολογικούς υποτύπους.
Γενετικός έλεγχος
H τεχνογνωσία σήμερα καλύπτει το πλέον ευρύ φάσμα γενετικού ελέγχου νεοπλασμάτων με μεθοδολογία και τεχνικές που είναι απαραίτητες για πολυποίκιλες προσεγγίσεις: PCR, real-time PCR (ποσοτική), ανάλυση αλληλουχίας των βάσεων DNA, blots, φθορίζων υβριδισμός in situ (FISH), multicolor FISH και CGH.
Η διαγνωστική αξία των χρωμοσωμικών ανακατατάξεων που περιλαμβάνονται στον πίνακα αναλύσεων που ακολουθεί είναι μεγάλη, καθώς και η δυνατότητα παρακολούθησης, σε ορισμένες περιπτώσεις, της υπολειπόμενης νόσου με real-time PCR [17, 18]
Γενετικές αναλύσεις σε αιματολογικά νεοπλάσματα
| Χρωμοσωμική Aνωμαλία | Υβριδικό Γονιδίο | Μεθοδολογία |
CML | t(9;22) | BCR/ABL | RT-PCR ή real-time PCR |
AML | t(8:21) | AML1/ETO | RT-PCR ή real-time PCR |
AML | t(15;17) | PML/RARA | RT-PCR ή real-time PCR |
AML | inv(16) | CBFB/MYH1 | RT-PCR |
ALL | SIL/TAL1 | RT-PCR | |
ALL | t(1;19) | E2A/PBX1 | RT-PCR |
ALL | t(4;11) | MLL/AF4 | RT-PCR |
ALL | t(9;22) | BCR/ABL | RT-PCR |
ALL | t(12;21) | TEL/AML1 | RT-PCR |
Επεξηγήσεις: CML: Χρόνια Μυελογενής Λευχαιμία, ΑΜL: Οξεία Μυελογενής Λευχαιμία, ALL: Οξεία Λεμφοβλαστική Λευχαιμία, RT-PCR: reverse transcription-PCR, t: μετάθεση, inv: αναστροφή, del:έλλειψη
Τα τελευταία λίγα χρόνια πολύ γνώση έχει αποκτηθεί στη μοριακή βάση από το διάφορες αναταραχές των υπεροξειδιοσωμάτων όπως συνοψίζεται κατωτέρω:
· Υπεροξειδιοσωμάτων βιογένεσης αναταραχές: μετά από τον προσδιορισμό από την εκτενή γενετική ετερογένεια μεταξύ των ασθενών που προσβλήθηκαν από από το PBD μέσα από σημπληρωματικές μελέτες, πολλά από να κρυμμένα γονίδια προσδιορίστηκαν και η μοριακή διάγνωση είναι τώρα δυνατή στη μεγάλη πλειοψηφία των περιπτώσεων.
· Rhizomelic chondrodysplasia punctata: τα τρία γονίδια που βρίσκονται πίσω από τους RCDP τύπους 1, 2 και 3 έχουν προσδιοριστεί και η μοριακή ανάλυση είναι τώρα δυνατή και για τις 3 αυτές μορφές.
· Υπεροξειδιοσωμάτων λιπαρού οξέος β-οξειδάση ανεπάρκειες: Acyl-CoA οξειδάσης ανεπάρκεια και D- δισλειτουργικής πρωτεϊνης ανεπάρκεια έχουν και οι δύο επιλυθεί σε μοριακό επίπεδο και πολύ συχνά ιδιαίτερες μεταλλαγές έχουν προσδιοριστεί. Το ίδιο πράγμα ισχύει για ασθένεια Refsum και την 2-methylacyl-CoA racemase ανεπάρκεια.[19]
Τα καρδιαγγειακά νοσήματα αποτελούν την πρώτη αιτία θανάτου στις αναπτυσσόμενες χώρες. Ο βιοχημικός προσδιορισμός δεικτών ελέγχου του καρδιαγγειακού συστήματος στον ορό ή το πλάσμα προσφέρει γρήγορη πληροφόρηση για τον κίνδυνο εμφάνισης καρδιαγγειακού νοσήματος. Σε συνδυασμό με τον βιοχημικό έλεγχο, αναπτύσσεται ταχύτατα και εφαρμόζεται πλέον ο γενετικός έλεγχος γονιδίων που σχετίζονται με τον κίνδυνο εμφάνισης καρδιαγγειακού νοσήματος. Η μοριακή ανάλυση τέτοιων γονιδίων μέσω ανίχνευσης συγκεκριμένων μεταλλάξεων προσφέρει τα παρακάτω οφέλη:
· Γίνεται μία φορά στην διάρκεια ζωής του ατόμου (lifetime test) προσφέροντας απόλυτα αξιόπιστα αποτελέσματα
· Προσφέρει προσυμπτωματική διάγνωση, δίνοντας την δυνατότητα πρόληψης και αυξημένης ιατρικής παρακολούθησης σε περίπτωση προσδιορισμού γονιδιακής μετάλλαξης που αυξάνει τον κίνδυνο εμφάνισης καρδιαγγειακών νοσημάτων. Το γεγονός αυτό έχει ιδιαίτερη σημασία στην περίπτωση νεογνών ή παιδιών μικρής ηλικίας, καθώς η θεραπευτική τους αγωγή ή η αγωγή πρόληψής τους μπορεί να ξεκινήσει το νωρίτερα δυνατόν.
· Σε κάποιες περιπτώσεις η ανίχνευση συγκεκριμένων γονιδιακών μεταλλάξεων οδηγεί σε αλλαγή θεραπευτικών σχημάτων προς όφελος του ασθενούς.
· Το αποτέλεσμα της γενετικής εξέτασης προσφέρει χρήσιμη ιατρική πληροφορία τόσο στα μέλη μιας οικογένειας, όσο και στους κοντινούς συγγενείς ενός εξεταζόμενου, καθώς στην περίπτωση ενός φορέα γονιδιακής μετάλλαξης, υπάρχει πιθανότητα, η μετάλλαξη αυτή να υπάρχει σε κάποιον από τους γονείς του, τα αδέλφια του ή τα παιδιά του.
Η ανίχνευση των κύριων γονιδιακών μεταλλάξεων που σχετίζονται με τα καρδιαγγειακά νοσήματα σε μια ομάδα διαφορετικών γονιδίων γίνεται μέσω προσδιορισμού νουκλεοτιδικής αλληλουχίας (DNA Sequencing). Η μέθοδος αυτή εντοπισμού μεταλλάξεων είναι απόλυτα ασφαλής και πλεονεκτεί έναντι όλων των υπόλοιπων μεθόδων (π.χ πέψη με περιοριστικά ένζυμα) στο θέμα της αξιοπιστίας αποτελεσμάτων.Σήμερα διεξάγεται γενετικός έλεγχος για τα κάτωθι:
· FACTOR V LEIDEN (Παράγων V LEIDEN)
· PROTHROMBIN (Προθρομβίνη)
· MTHFR (Γονίδιο Υπερομοκυστεϊναιμίας)
· CBS (β-Συνθετάση της Κυσταθειονίνης)
· Glycoprotein IIIa (Γλυκοπρωτεΐνη ΙΙΙα)
· eNOS (Endothelial Nitric Oxide Synthase) (Ενδοθηλιακή Συνθετάση του Νιτρικού Οξειδίου)
· Stromelysin-1(matrix metalloproteinase-3) (Στρωμελυσίνη-1)
· Apolipoprotein E (Απολιποπρωτεΐνη Ε)
· Alpha-1-antitrypsin (Α1ΑΤ) (Άλφα-1 Αντιτρυψίνη )
· ACE (Ένζυμο Μετατροπής Αγγεοιοτενσίνης) [20]
FACTOR V LEIDEN (Παράγων V LEIDEN)
Η αντοχή (resistance) στην ενεργοποιημένη πρωτεΐνη C είναι η πιο συχνή αιτία φλεβικής θρόμβωσης. Η αιτία της φλεβικής θρόμβωσης συνδέεται σε μεγάλο ποσοστό με την ύπαρξη μιας συγκεκριμένης μετάλλαξης στο γονίδιο FACTOR V. Η μετάλλαξη αυτή έχει πλέον χαρακτηρισθεί (G1691A) και αφορά την αντικατάσταση ενός αμινοξέος από κάποιο άλλο. Στο γενικό πληθυσμό περίπου το 6-10% των αντρών και γυναικών Καυκάσιας καταγωγής είναι ετερόζυγοι για τη μετάλλαξη αυτή, ενώ πιο σπάνια είναι η περίπτωση ομόζυγων ατόμων. Η ύπαρξη της μετάλλαξης στο γονίδιο του παράγοντα V Leiden αυξάνει την πιθανότητα εμφάνισης θρόμβωσης κατά 7 φορές στους ετεροζυγώτες και κατά 80 φορές στους ομοζυγώτες. Η ανίχνευση της μετάλλαξης αιτιολογεί την εμφάνιση θρόμβωσης, αποκαλύπτει άτομα και οικογένειες με υψηλές πιθανότητες εμφάνισης θρόμβωσης και παρέχει την δυνατότητα λήψης προληπτικών μέτρων για άτομα που έχουν υψηλότερο κίνδυνο εμφάνισης φλεβικών θρομβώσεων.
Ανίχνευση της μετάλλαξης G1691A του γονιδίου Factor V [20]
PROTHROMBIN (Προθρομβίνη)
Η προθρομβίνη είναι ο πρόδρομος της πρωτεάσης θρομβίνης, η οποία έχει τον κύριο ρόλο στη διαδικασία της αιμόστασης και της θρόμβωσης. Είναι πλέον αποδεκτό ότι η νουκλεοτιδική αντικατάσταση μιας βάσης G σε A στη θέση 20210 (G20210Α) συνδέεται άμεσα με τη φλεβική θρόμβωση. Οι φορείς της συγκεκριμένης μετάλλαξης εμφανίζουν 2,8 φορές μεγαλύτερο κίνδυνο εμφάνισης θρόμβωσης. Επομένως, κρίνεται αναγκαία η ανίχνευση της μετάλλαξης στο γονίδιο της προθρομβίνης.
Ανίχνευση της μετάλλαξης G20210Α του γονιδίου της προθρομβίνης [20]
MTHFR (Γονίδιο Υπερομοκυστεϊναιμίας)
Η υπερομοκυστεϊναιμία είναι ένας κύριος παράγοντας εμφάνισης μεταξύ άλλων ασθενειών και της φλεβικής θρόμβωσης. Τα επίπεδα της ομοκυστεΐνης στο αίμα επηρεάζονται τόσο από γενετικούς όσο και από περιβαλλοντικούς παράγοντες. Ένας από τους κύριος γενετικούς παράγοντες είναι η ύπαρξη μεταλλάξεων στο γονίδιο MTHFR. Σήμερα, είναι καλά χαρακτηρισμένες δύο μεταλλάξεις στο συγκεκριμένο γονίδιο : η C677T και η A1298C. Η πρώτη από αυτές έχει συνδεθεί άμεσα με υπερομοκυστεϊναιμία, τόσο σε ομόζυγη όσο και σε ετερόζυγη κατάσταση. Αντίθετα, η δεύτερη δεν έχει συσχετισθεί με υψηλά επίπεδα ομοκυστεΐνης στο αίμα. Τέλος, η ταυτόχρονη ύπαρξη και των δύο μεταλλάξεων σε ετερόζυγη κατάσταση έχει τα ίδια συμπτώματα με τη μετάλλαξη C677T σε ομόζυγη κατάσταση. Επομένως, η ανίχνευση μεταλλάξεων στο γονίδιο MTHFR είναι σημαντική τόσο για τον προσδιορισμό οικογενειών με αυξημένη πιθανότητα φλεβικής θρόμβωσης, όσο και για το σχεδιασμό κατάλληλης θεραπευτικής αγωγής.
Ανίχνευση των μεταλλάξεων C677T και A1298C του γονιδίου MTHFR.[20]
CBS (β-Συνθετάση της Κυσταθειονίνης)
Το ένζυμο CBS μετατρέπει την ομοκυστεΐνη σε κυσταθειόνη. Οι μεταλλάξεις του γονιδίου αυτού έχουν ως αποτέλεσμα την παραγωγή πρωτεΐνης μειωμένης ενεργότητας, με αποτέλεσμα την αύξηση των επιπέδων της ομοκυστεΐνης στο αίμα. Μέχρι σήμερα, έχουν περιγραφεί 131 μεταλλάξεις στο συγκεκριμένο γονίδιο, οι οποίες εντοπίζονται σε όλο το μήκος του και οι οποίες σχετίζονται με συγκεκριμένα κλινικά ευρήματα. Οι πιο βασικές είναι η G919A και η T833C.
Ανίχνευση των μεταλλάξεων G919A και T833C του γονιδίου CBS.[20]
PAI-1 (Ενεργοποιητής του Aναστολέα του Πλασμινογόνου)
Ο παράγων Plasminogen Activator Inhibitor-1 (PAI-1) είναι ο βασικός αναστολέας της ινοδώλυσης. Όταν τα επίπεδα του PAI-1 είναι υψηλά τότε η ινοδώλυση αναστέλλεται και αυξάνει ο κίνδυνος για αρτηριακή ή φλεβική θρόμβωση. Έχει αναγνωρισθεί μια μετάλλαξη στο γονίδιο PAI-1, η οποία έχει ως αποτέλεσμα την ύπαρξη δύο αλληλομόρφων του γονιδίου : 4G και 5G. Η παρουσία των 4G/4G και 4G/5G γονοτύπων στον πληθυσμό ανέρχεται στο 28% και 48% αντίστοιχα, και οι συγκεκριμένοι γονότυποι έχουν συσχετισθεί άμεσα με υψηλά επίπεδα ενεργότητας του PAI-1 στο πλάσμα και κατά συνέπεια με αυξημένη πιθανότητα εμφάνισης καρδιαγγειακών παθήσεων και εμφραγμάτων. Επομένως, είναι αναγκαία η ανίχνευση των αλληλομόρφων του ατόμου, ώστε να λάβει την απαραίτητη ιατρική παρακολούθηση.
Ανίχνευση των αλληλομόρφων 4G και 5G στο εξεταζόμενο άτομο.[20]
Glycoprotein IIIa (Γλυκοπρωτεΐνη ΙΙΙα)
Η Glycoprotein IIIa είναι μέρος ενός υποδοχέα που βρίσκεται στην επιφάνεια των αιμοπεταλίων και διαμεσολαβεί στην διαδικασία συσσωμάτωσης των αιμοπεταλίων, διαδραματίζοντας σημαντικό ρόλο στην αιμόσταση. Έχουν περιγραφεί δυο πολυμορφισμοί του γονίδιου (P1A1, P1A2). Η συχνότητα εμφάνισης τους στο πληθυσμό είναι 64%, 34% και 2,4% για τους γονοτύπους P1A1/P1A1, P1A1/P1A2 και P1A2/P1A2 αντίστοιχα. Ο πολυμορφισμός P1A2 έχει σχετισθεί άμεσα με περιπτώσεις εμφράγματος του μυοκαρδίου σε πρώιμες ηλικίες (<55 ετών), καθώς και με αυξημένη πιθανότητα θρόμβωσης της στεφανιαίας αρτηρίας σε ασθενείς που έχουν υποστεί καρδιολογικές επεμβάσεις με την χρήση stent.Επομένως, κρίνεται επιτακτική ανάγκη ο προσδιορισμός του γονοτύπου ατόμων με καρδιαγγειακά προβλήματα.
Ανίχνευση των πολυμορφισμών P1A1 και P1A2 του γονιδίου Glycoprotein IIIa.[20]
eNOS (Endothelial Nitric Oxide Synthase) (Ενδοθηλιακή Συνθετάση του Νιτρικού Οξειδίου)
Το ένζυμο eNOS καταλύει την σύνθεση του ΝΟ, το οποίο παίζει σημαντικό ρόλο στην διατήρηση του αγγειακού τόνου. Η οποιαδήποτε ανεπάρκεια στη σύνθεση του ΝΟ αυξάνει τον κίνδυνο για πολύ σοβαρές καρδιαγγειακές δυσλειτουργίες (σύσπαση στεφανιαίας αρτηρίας, ατελές κάταγμα μυοκαρδίου κ.α.). Η παρουσία μεταλλάξεων στο γονίδιο του eNOS (T786C) έχει ως αποτέλεσμα την μειωμένη σύνθεση του ΝΟ και κατά συνέπεια την αύξηση του κινδύνου για τις προαναφερόμενες καρδιαγγειακές δυσλειτουργίες. Έχουν ανιχνευθεί δύο μεταλλάξεις στο γονίδιο eNOS οι οποίες συνδέονται άμεσα με τις προαναφερόμενες παθήσεις. Η μετάλλαξη G894T (Glu298Asp) και η μετάλλαξη T786C, οι οποίες οδηγούν σε μειωμένη σύνθεση ΝΟ και αυξάνουν την πιθανότητα καρδιαγγειακών παθήσεων. Αν λάβουμε, επίσης, υπόψη μας τη δυσκολία διάγνωσης της σύσπασης στεφανιαίας αρτηρίας, κρίνεται επιτακτική ανάγκη η γενετική εξέταση οικογενειών με αυξημένη πιθανότητα εμφάνισης της συγκεκριμένης πάθησης.
Ανίχνευση των μεταλλάξεων G894T και T786C του γονιδίου eNOS.[20]
Stromelysin-1(matrix metalloproteinase-3) (Στρωμελυσίνη-1)
Η Stromelysin-1 είναι ένα ένζυμο που δρα στο εξωκυττάριο πλέγμα συμμετέχοντας στη διαδικασίες επαναδημιουργίας του. Η σύσταση του εξωκυττάριου πλέγματος παίζει κρίσιμο ρόλο στην συσσώρευση λιπιδίων, την αναδημιουργία των αγγείων και την σταθερότητα της αθηρωματικής πλάκας. Η παρουσία ενός πολυμορφισμού (6A) στο γονίδιο της Stromelysin-1, ακόμη και σε ετερόζυγη κατάσταση, σχετίζεται με χαμηλά επίπεδα του ενζύμου και αυξημένη ευαισθησία σε παράγοντες που συμβάλουν στην εμφάνιση των καρδιαγγειακών νοσημάτων. Η συχνότητα αυτού του αλληλόμορφου στο γενικό πληθυσμό είναι μεγάλη (30%) και η ανίχνευση της παρουσίας του θα μπορούσε να οδηγήσει σε λήψη μέτρων (αλλαγές τρόπου ζωής) που να μειώνουν τον κίνδυνο εμφάνισης καρδιαγγειακών νοσημάτων.
Ανίχνευση των πολυμορφισμών 5Α και 6A στο εξεταζόμενο άτομο [20]
Apolipoprotein E (Απολιποπρωτεΐνη Ε)
Η εξέταση αυτή αφορά την μελέτη του γονιδίου της Απολιποπρωτείνης Ε ως προς τον προσδιορισμό αλληλομόρφων τα οποία μπορούν να ευθύνονται για:
-Αθηρωματική στένωση των αιμοφόρων αγγείων, έμφραγμα μυοκαρδίου
-Αυξημένα ποσά χοληστερόλης και β-λιποπρωτείνης
-Οικογενή υπερλιπιδαιμία τύπου ΙΙΙ
Υπάρχουν τρία αλληλόμορφα (e2, e3, e4) του γονίδίου της απολιπωπρωτείνης Ε, τα οποία κωδικοποιούν τις τρεις ισομορφές της πρωτεΐνης apoE. Από αυτά τα e2 και e4 έχουν συνδεθεί άμεσα με υψηλές συγκεντρώσεις τριγλυκεριδίων στο αίμα.
Ο προσδιορισμός των αλληλομόρφων της Απολιποπρωτεΐνης Ε με αλληλούχιση ειδικών περιοχών του γονιδίου είναι μια ταχεία και απόλυτα ακριβής μέθοδος με χαμηλό κόστος ανάλυσης που πλεονεκτεί έναντι των υπολοίπων στο θέμα της αξιοπιστίας. Η παραπάνω εξέταση προτείνεται σε όλους τους ασθενείς που εμφανίζουν αυξημένα επίπεδα ολικής χοληστερόλης (>260 mg/ml) παράλληλα με αυξημένα επίπεδα τριγλυκεριδίων (>300 mg/ml).
Ανίχνευση των αλληλομόρφων e2, e3 και e4 στο εξεταζόμενο άτομο [20]
Alpha-1-antitrypsin (Α1ΑΤ) (Άλφα-1 Αντιτρυψίνη)
Η άλφα-1-Αντιτρυψίνη προστατεύει τους συνδετικούς ιστούς των πνευμόνων από ένα φυσικό ένζυμο (neutrophil elastase), ενώ η καταστολή της είναι συχνή γενετική αιτία για αυξημένη πιθανότητα κίρρωσης και εμφυσήματος. Έχουν ανιχνευθεί δύο μεταλλάξεις, S και Z, στο γονίδιο A1AT, οι οποίες ευθύνονται για μειωμένη δράση της A1AT πρωτεΐνης. Ειδικότερα, η Ζ μετάλλαξη αφορά μια νουκλεοτιδική αντικατάσταση στο κωδικόνιο 342 του γονιδίου, ενώ η S μετάλλαξη μια αντίστοιχη, στο κωδικόνιο 264. Έχει διαπιστωθεί ότι η ομοζυγωτία για την Ζ μετάλλαξη ή η παράλληλη ετεροζυγωτία για την Ζ και την S μετάλλαξη οδηγούν σε εμφύσημα σε άτομα νεαρής ηλικίας καθώς και σε ασθένειες του ήπατος τόσο σε νεογνά όσο και σε ενήλικες.
Ο προσδιορισμός των συγκεκριμένων μεταλλάξεων του γονιδίων με αλληλούχιση ειδικών περιοχών του είναι μια ταχεία και απόλυτα ακριβής μέθοδο με χαμηλό κόστος ανάλυσης που πλεονεκτεί έναντι των υπολοίπων στο θέμα της αξιοπιστίας. Ανίχνευση των μεταλλάξεων S και Z του γονιδίου Α1ΑΤ.[20]
ACE (Ένζυμο Μετατροπής Αγγειοτενσίνης)
Το ένζυμο ACE διαμεσολαβεί στο μεταβολισμό της αγγειοτενσίνης και τα επίπεδα του σχετίζονται με την υπέρταση, τη διαβητική νεφροπάθεια και με καρδιαγγειακά νοσήματα. Τα επίπεδα του ένζυμου συνδέονται στενά με την παρουσία ενός πολυμορφισμού (Insertion/Deletion polymorphism) στο ιντρόνιο 16 του γονιδίου ACE και επομένως ο έλεγχος για την παρουσία του πολυμορφισμού μπορεί να συμβάλει στην πληρέστερη διάγνωση και πρόληψη των σχετιζόμενων νοσημάτων. Ανίχνευση του πολυμορφισμού του γονιδίου ACE.[20]
[1] Ken Mills, Methods in Molecular Medicine, vol. 68: Molecular Analysis of Cancer, Edited by: J. Boultwood and C. Fidler, Humana Press Inc.,
[2] Γούσια Α., M.D., Ph.D, Σύγχρονες απόψεις για την αξιολόγηση των καρκινικών δεικτών στη χειρουργική, Εργαστήριο Παθολογικής Ανατομικής, Ιατρική Σχολή Πανεπιστημίου Ιωαννίνων.
[3] M van de Rijn & C B Gilks, Histopathology, 44:97-108, 2004
[4] ΤΣΑΚΟΥΝΤΑΚΗΣ Ν., ΤΑ ΕΠΙΔΗΜΙΟΛΟΓΙΚΑ ΔΕΔΟΜΕΝΑ ΤΟΥ ΚΑΡΚΙΝΟΥ ΤΟΥ ΜΑΣΤΟΥ ΣΤΗ ΚΡΗΤΗ ΚΑΙ Η ΣΥΣΧΕΤΙΣΗ ΤΟΥΣ ΜΕ ΠΡΟΓΝΩΣΤΙΚΟΥΣ ΠΑΡΑΓΟΝΤΕΣ, Διδακτορική Διατριβή, ΗΡΑΚΛΕΙΟ ΚΡΗΤΗΣ 2005
[5] Kalomenidis I, Orphanidou D, Papamichalis G, Vassilakopoulos T, Skorilas A, Rasidakis A, Papastamatiou H, Jordanoglou J, Roussos C.,Combined expression of p53, Bcl-2, and p21WAF-1 proteins in lung cancer and premalignant lesions: association with clinical characteristics,Department of Pulmonary and Critical Care Medicine, Medical School of Athens University, Evangelismos Hospital, Athens, Greece, The Forum for Early Diagnosis and Treatment of Lung Cancer, Number 14 December 2002.
[6] Σ. ΠΑΠΑΔΟΠΟΥΛΟΥ, Α. ΣΚΟΡΙΛΑΣ, Ν. ΑΡΝΟΓΙΑΝΝΑΚΗ, Ι. ΓΙΩΤΗΣ, Β. ΠΑΠΑΠΑΝΑΓΙΩΤΟΥ, Ν. ΑΓΝΑΝΤΗ, Μ. ΤΑΛΙΕΡΗ, Ο ΕΝΕΡΓΟΠΟΙΗΤΗΣ ΤΟΥ ΠΛΑΣΜΙΝΟΓΟΝΟΥ UΡΑ. ΕΝΑΣ ΝΕΟΣ ΔΕΙΚΤΗΣ ΓΙΑ ΤΟΝ ΚΑΡΚΙΝΟ ΤΟΥ ΠΑΧΕΟΣ ΕΝΤΕΡΟΥ, ΙΑΤΡΙΚΗ, 78(5), 451-456, 2000 - Ερευνητική εργασία.
[7] Α. Γκέκας, Α.Γ. Παπατσώρης, Ι. Δάρρας, ΟΓΚΟΛΟΓΙΚΟΙ ΔΕΙΚΤΕΣ ΣΤΟΝ ΚΑΡΚΙΝΟ ΤΟΥ ΟΥΡΟΠΟΙΗΤΙΚΟΥ, Ουρολογική Κλινική Γ.Π.Ν. Πατρών, http://www.oncology.gr/1/synedrio1/praktikaa17.HTM
[8] Παν. Ν. Γεωργακόπουλος, ΟΓΚΟΛΟΓΙΚΟΙ ΔΕΙΚΤΕΣ ΣΤΟΝ ΚΑΡΚΙΝΟ ΤΟΥ ΠΝΕΥΜΟΝΑ, ΜΕΘ ΠΓΝΠ "Αγιος Ανδρέας", http://www.oncology.gr/1/synedrio1/praktikaa16.HTM
[9] Πέτρου Παναγιώτα, ΣΥΣΤΟΙΧΙΕΣ ΜΙΚΡΟΚΗΛΙ∆ΩΝ ΟΛΙΓΟΝΟΥΚΛΕΟΤΙ∆ΙΩΝ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΜΕΤΑΛΛΑΞΕΩΝ, Εργαστήριο Ανοσοαναλύσεων/Ανοσοαισθητήρων, Ινστιτούτο Ραδιοϊσοτόπων ––Ραδιοδιαγνωστικών Προϊόντων, ΕΚΕΦΕ «∆ηµόκριτος», Θερινό σχολείο, 11-22 Ιουλίου, 2005.
[11] Knutila 2004 Ann Med 36(3):162-71
[12] Pandis 1999 Forum Clin Oncol 2(4):293-8
[13] Seidl et al. 2003 Lancet Oncol 4(9):557-64
[14] Angtuaco et al 2004 Radiology 1: 11-23
[15] Nowell & Hungerford 1960 Science 132: 1497
[16] Mitelman et al 2004 http://cgap.nci.nih.gov/Chromosomes/Mitelman
[17] van Dongen et al 2003 Leukemia 13: 1901-28
[18] Gabert et al. 2003 Leukemia 17: 2318-57
[19] Nenad Blau, et all, Physician’s Guide to the laborator Diagnosis of Metabolic Diseases, Spinger 2ed ed. 2003, p.506.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
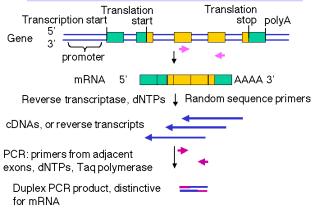{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Δεν υπάρχουν σχόλια:
Δημοσίευση σχολίου